Понимание значения этого большого и незаменимого органа пока еще находится в жалком состоянии. Частично это объясняется тем, что он очень сложен, не только структурно, но и функционально, а частично тем, что интуитивные предположения нейробиологов относительно его функций весьма часто оказывались неверными. Однако сейчас перспектива изменяется в связи с тем, что методы усовершенствовались, а исследователи научились тому, как обращаться с огромным числом охваченных сложнейшими связями нейронов, которые являются основными элементами коры, с импульсами, которые они проводят, и с синапсами, с помощью которых они связаны. В настоящей статье мы попытаемся обрисовать современное состояние знаний, касающихся одной из подобластей коры - первичной зрительной проекционной зоны (известной также под названием стриарной коры, или поля 17) - самой простой из тех кортикальных зон, которые связаны со зрением. Это неизбежно приведет нас к рассмотрению соответствующих вопросов зрительного восприятия, так как работу какого-либо органа нелегко отделить от его биологического назначения.
Кора большого мозга - сильно складчатая нервная ткань толщиной около двух миллиметров - представляет собой самую наружную оболочку, покрывающую верхнюю часть полушарий головного мозга и несколько заходящую на их нижнюю поверхность. У человека вся площадь коры, если ее разгладить, равна примерно полутора квадратным футам. (В 1963 г. в своей статье в «Scientific American» один из нас привел для этой площади цифру 20 квадратных футов и был тут же поправлен нашим другом нейроанатомом из Торонто, который сказал, что, по его мнению, эта площадь составляет около 1,5 квадратных фута, «по крайней мере у канадцев».) Складчатость, по-видимому, в основном является результатом того, что такую необычную структуру нужно было упаковать в объеме черепа.
При рассмотрении коры под микроскопом сразу же бросается в глаза огромное множество нейронов: около 105 (100000) на каждый квадратный миллиметр поверхности, откуда можно заключить, что вся кора в целом содержит что-то около 1010 (10 миллиардов) нейронов. Тела клеток образуют полдюжины слоев, причем слои с плотным и редким расположением клеток чередуются. В противоположность столь выраженным различиям в плотности клеток в последовательных слоях коры на различной глубине в плоскости каждого данного слоя распределение явно однородно во всех участках и по всем направлениям. Морфологически кора довольно однородна по двум своим измерениям.
Одно из первых великих прозрений, касающихся организации коры, относится к концу XIX в., когда постепенно осознали, что этот сравнительно однородный пласт ткани подразделяется на некоторое число отдельных областей, сильно различающихся по функции. Свидетельства в пользу этого поступали из клинических, физиологических и анатомических источников. Было замечено, что повреждения мозга, в зависимости от их локализации, могут вызывать либо паралич, либо слепоту, либо потерю тактильной чувствительности, либо потерю речи; при этом слепота бывает общей или проявляющейся только в половине или меньшей части поля зрения, а потеря чувствительности может затрагивать одну конечность или несколько пальцев. Устойчивость взаимосвязи между данным дефектом и локализацией данного поражения позволила постепенно составить карты наиболее очевидных из этих специализированных областей: зрительной, слуховой, соматосенсорной (ощущение тела), речевой и моторной.
Во многих случаях более внимательное рассмотрение под микроскопом коры, окрашенной для выявления клеточных тел, показывает, что, несмотря на относительную однородность, там имеются структурные вариации, особенно в характере слоистости, которые хорошо коррелируют с клинически выявленными подразделениями. Дополнительное подтверждение этого последовало из наблюдений, касающихся локализации (на поверхности мозга) электрических волн, возникающих при стимуляции животного прикосновением к телу, звуковыми щелчками или тональными сигналами, действующими на ухо, или вспышками света перед глазом. Сходным образом, моторные области могут быть картированы с помощью электрической стимуляции коры и регистрации того, какая часть тела животного при этом приходит в движение. Такое систематическое картирование коры вскоре привело к фундаментальному заключению: большинство сенсорных и моторных областей содержит закономерно построенные систематические двумерные карты того внешнего мира, который они представляют. Разрушение некоторого небольшого участка коры может привести к параличу одной руки; сходное повреждение в другой ограниченной области приводит к «онемению» одной кисти или верхней губы, или к слепоте, проявляющейся в небольшой части поля зрения; если на кору мозга животного поместить электроды, можно установить, что прикосновение к одной конечности вызывает серию соответственным образом локализованных электрических потенциалов. Несомненно, что тело представлено систематическим образом в соматосенсорной и моторной зонах, а зрительный мир представлен в виде карты в первичной зрительной коре-области затылочной доли, которая у человека и обезьян-макаков (того животного, на котором в основном проводились наши исследования) имеет площадь около 15 квадратных сантиметров.
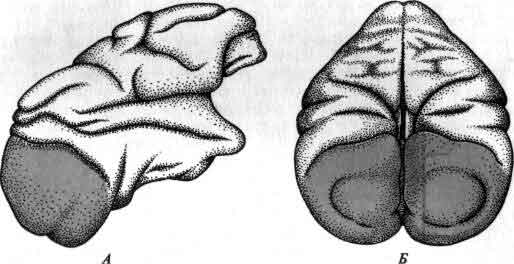
Первичная зрительная кора, известная также под названием стриaрной коры, или поля 17, - участок коры большого мозга, слоистой нейронной ткани, покрывающей мозг приматов. В мозгу макака, для которого здесь показаны вид сбоку (А) и сверху и сзади (Б), первичная зрительная кора (окрашенная область) занимает большую часть видимой поверхности обеих затылочных долей. Она также загибается на медиальную поверхность мозга. Кроме того, она продолжается в виде сложных складок и под выпуклой наружной поверхностью, как это показано на парасаггитальном срезе (см. следующий рисунок), который был сделан вдоль цветной линии и рассматривается со стороны, указанной стрелками.
В первичной зрительной коре карта не осложнена разрывами и смещениями; наблюдается лишь примечательное расщепление мира точно по его середине, причем левая его половина проецируется на правую часть мозговой коры, а правая половина — на левую. Карта тела более сложна и, по всей вероятности, пока еще не до конца выяснена. Тем не менее в ней есть своя система и она сходным образом перекрещена: правая часть тела проецируется в левое полушарие, а левая часть — в правое. (Следует заметить, что ни у кого нет даже самой отделенной идеи относительно того, какой смысл может заключаться в этой удивительной тенденции к перекрещиванию нервных путей.)
Важной особенностью кортикальных карт является наличие в них искажений. Масштабы этих карт варьируют наподобие проекций Меркатора, причем правило для коры такое: области наибольшего разрешения или тонкости функций занимают относительно большие области коры. Если взять поверхность тела, то миллиметр поверхности пальцев, губ или языка проецируется на большую часть коры, чем миллиметр туловища, ягодиц или спины; в зрении центральная часть сетчатки представлена примерно в 35 раз более детально, чем периферические области.
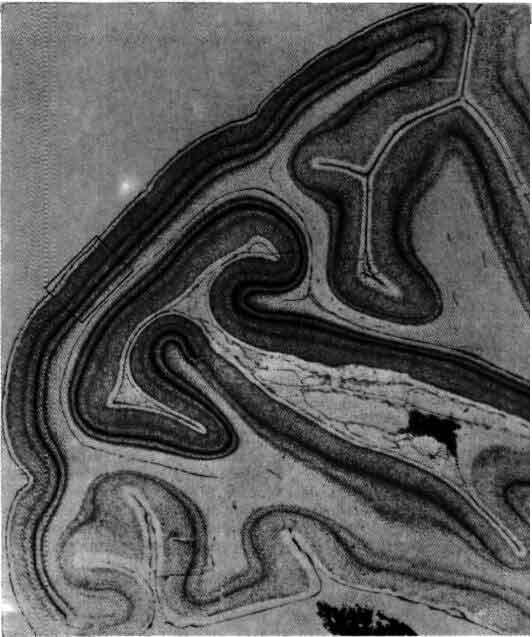
Срез зрительной коры, прошедший вдоль цветной линии, показанной на предыдущем рисунке, окрашен по методу Ниссля, выявляющему тела клеток, но не волокна. Видно, что зрительная кора представляет собой непрерывный слой нейронов толщиной около 2 мм. В рамке участок среза, показанный при большем увеличении на следующем рисунке.
Существенно, что достижения в составлении карт кортикальных проекций иногда уводили в сторону от главной проблемы: как мозг анализирует информацию. Получалось так, будто установление представительства само по себе может быть конечным итогом, а не служит более скромной цели, — как если бы задачей коры было снабжение информацией маленького гномика, который сидит внутри головы и рассматривает образы, возникающие на коре. В ходе этой статьи мы покажем, что по крайней мере в зрении мир представлен в гораздо более искаженном виде; любой маленький гномик, пытающийся собрать по частям информацию с кортикальных проекций, пришел бы в крайнее замешательство.

Поперечное сечение первичной зрительной коры макака (окраска по методу Ниссля) демонстрирует слоистую структуру и представляет общепринятые обозначения шести слоев. Белые пустоты - срезы кровеносных сосудов.
Тем не менее первым важным прозрением относительно организации коры было именно осознание этого разделения на зоны, имеющие сильно различающиеся функции и тенденцию к упорядоченной планировке. Сколько же точно имеется таких зон — этот вопрос был предметом широких спекуляций. Оценки анатомов в целом весьма высоки и доходят до нескольких сотен зон; конкретные цифры зависят от индивидуальной чувствительности исследователя к тонким различиям микроскопических структур, а иногда также и от его способности обманываться. Физиологи начали с более низких оценок, но позже, в результате использования более эффективных методов картирования, пересмотрели эти оценки в сторону их повышения. Важная основополагающая идея состоит в том, что информация любой данной модальности, например зрительная или слуховая, поступает сначала в первичную зону коры, а оттуда, либо прямым путем, либо через таламус, последовательно передается в ряд высших центров. Современные догадки относительно числа зон коры колеблются между 50 и 100.
Второе важное прозрение относительно организации коры связано с работами анатома С. Рамон-и-Кахала и его ученика Р. Лоренте де Но. Это было осознание того, что операции, которые кора осуществляет над получаемой информацией, локальны. Что это означает, легче всего понять, рассматривая схему связей, установленную с помощью метода Гольджи, которой пользовались Рамон-и-Кахал и Лоренте де Но. В сущности, связи эти просты. Пучки волокон несут информацию в кору; на пути они прерываются несколькими синаптическими переключениями и вводят информацию во все клеточные слои, пересекаемые по вертикали; наконец, по нескольким другим пучкам волокон модифицированные сообщения выходят из данной зоны коры. Детали связей между входами и выходами различаются от зоны к зоне, но внутри данного поля они кажутся довольно стереотипными. Общим для всех зон является локальный характер связен. Информация, поступающая в кору по одному волокну, может в принципе пройти сквозь всю толщину коры, примерно через три или четыре синапса, тогда как расползание в стороны, производимое ветвлениями аксонов и дендритов, практически во всех случаях ограничено несколькими миллиметрами - малой частью обширной площади коры.
Это имеет далеко идущие следствия. Что бы данная область коры ни делала, она делает это локально. На стадиях, где имеет место детальное систематическое топографическое картирование любого типа, анализ должен быть «дробным». Так, например, в соматосенсорной коре сигналы от одного пальца могут комбинироваться и сопоставляться со входами от этого же или от соседнего пальца, но они едва ли будут комбинироваться с влияниями от туловища или от ноги. То же самое применимо и к зрительному миру. Если взять детальную схему входов в первичную зрительную кору, то покажется невероятным, чтобы эта область могла каким-нибудь способом сопоставить информацию, поступающую из точек, расположенных намного выше и намного ниже горизонта или из правой и левой половин рассматриваемой картины. Какие бы операции эти кортикальные зоны ни выполняли, это должен быть какого-то рода локальный анализ сенсорного мира. Можно лишь предположить, что по мере того, как зрительная, тактильная или слуховая информация передается от одной зоны коры к следующей, карты становятся все более и более расплывчатыми, а передаваемые сообщения - все более и более абстрактными.
Хотя исследования с применением метода Гольджи еще в начале девятисотых годов прояснили, что кора должна выполнять локальный анализ, прошло полстолетия, прежде чем у физиологов появились хоть какие-то идеи относительно того, какого же именно типа должен быть этот анализ в разных зонах коры. Первые открытия были сделаны на первичной зрительной зоне, являющейся сейчас наиболее изученной и пока еще единственной, в которой анализ и последовательные преобразования информации известны сколько-нибудь детально. После описания основных преобразований, которые имеют место в первичной зрительной коре, мы покажем, как успехи в понимании этих кортикальных функций открыли целый мир схем организации, который иначе оказался бы недоступным наблюдению.
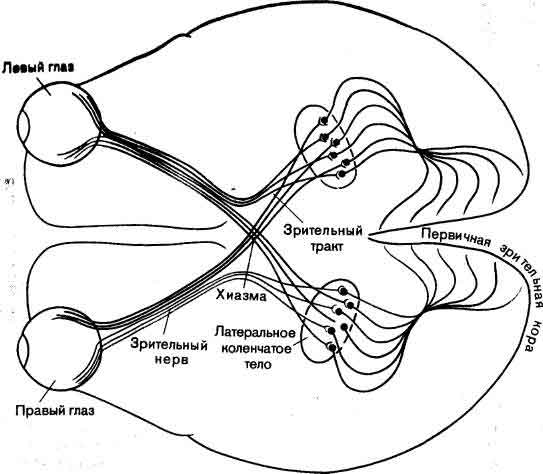
Схема зрительных путей мозга человека (вид снизу). По аксонам ганглиозных клеток, объединенным в зрительный нерв, выходные сигналы сетчатки передаются в латеральные коленчатые тела; примерно половина аксонов перекрещивается и идет на противоположную сторону мозга, так что представление каждой половины зрительной сцены проецируется на коленчатое тело противоположного полушария мозга. Нейроны коленчатых тел посылают свои аксоны в первичную зрительную кору.
Начать нам лучше всего с прослеживания зрительного пути у приматов от сетчатки до коры. Выходные сигналы каждого глаза идут в мозг примерно по миллиону нервных волокон, объединенных в зрительный нерв. Эти волокна — аксоны ганглиозных клеток сетчатки. Прежде чем сигналы светочувствительных элементов — палочек и колбочек — дошли до ганглиозных клеток, они уже преодолели от двух до четырех синапсов и вовлекли в работу четыре других типа клеток, так что довольно сложный анализ информации уже имел место.

Латеральное коленчатое тело обезьяны (А) - слоистая структура, в которой клетки слоев 1, 4 и 6 (если считать снизу вверх) получают на вход сигналы от контралатерального глаза, а слоев 2, 3 и 5 - от ипсилатерального. Карты расположены упорядоченно, так что нейроны, лежащие вдоль любого радиуса (черная линия), получают сигналы от одной и той же части зрительной картины. Послойное распределение входящих аксонов демонстрируется на двух латеральных коленчатых телах животного, у которого был зрячим только левый глаз (Б, В): в каждом коленчатом теле клетки трех слоев, имеющих входы от правого глаза, атрофированы.
Большая часть волокон зрительного нерва идет, не прерываясь, к двум клеточным ядрам, расположенным в глубине мозга; ядра эти называются латеральными коленчатыми телами; здесь волокна образуют синапсы. В свою очередь клетки латеральных коленчатых тел направляют свои аксоны прямо в первичную зрительную кору. Оттуда после нескольких синаптических переключений сообщения посылаются в ряд более далеких пунктов назначения: в соседние области коры, а также в несколько отделов в глубине мозга. Одна группа волокон даже направляется обратно в латеральные коленчатые тела; функция этой цепи обратной связи неизвестна. На данный момент самое главное то, что первичная зрительная кора ни в коей мере не является концом зрительного пути. Это только первая ступень, возможно, весьма ранняя в смысле степени абстрагирования информации, которую она обрабатывает.
Вследствие частичного перекреста зрительных нервов в хиазме коленчатое тело и кора левой стороны соединены с левыми половинами обеих сетчаток и, соответственно, имеют дело с правой половиной поля зрения, а для коленчатого тела и коры правой стороны справедливо обратное. Каждое коленчатое тело и кора получают сигналы от двух глаз и имеют дело с противоположной половиной зрительного мира.
Наша стратегия исследования работы этого зрительного пути с конца 50-х годов была в принципе простой. Начав, скажем, с волокон зрительного нерва, мы регистрировали микроэлектродами активность отдельных волокон и пытались найти способ наиболее эффективного влияния на их разряд путем стимуляции сетчатки светом. Для этой цели можно использовать световые стимулы любого мыслимого размера, формы и цвета, яркие на темном фоне и, наоборот, неподвижные и движущиеся. Хотя это порой отнимало много времени, но раньше или позже мы добивались того, что находили наилучший стимул для исследуемой клетки, в данном случае - ганглиозной клетки сетчатки. (Иногда мы ошибались!) Мы регистрировали результаты и переходили к другому волокну. После обследования нескольких сотен клеток могло обнаружиться, что новые типы попадаются редко. Удовлетворившись тем, что мы примерно знаем, как работают нейроны этой ступени, мы переходили к следующей ступени (в данном случае - коленчатому телу) и повторяли процедуру. Сравнение двух наборов результатов могло сказать нам кое-что о характере преобразований в коленчатом теле. Тогда мы шли далее к следующей ступени - первичной коре - и снова повторяли процедуру.
Действуя таким образом, удалось обнаружить, что ганглиозные клетки сетчатки и клетки коленчатого тела лучше всего реагируют на приблизительно круглые пятна некоторого конкретного размера в определенной части поля зрения. Размер является критическим параметром, поскольку рецептивное поле каждой клетки (совокупность рецепторов сетчатки, посылающих сигналы к данной клетке) целится на две части и имеет либо возбуждающий центр и тормозную периферию (клетки с «on-центром»), либо прямо противоположную организацию (клетки с «off-центром»). Такая концентрическая организация впервые была описана С. Куффлером (S. Kuffler) из Медицинской школы Университета Джонса Гопкинса в 1953 г. Пятно, в точности совпадающее с центром рецептивного поля, служит при этом более эффективным стимулом, чем пятно большего размера, заходящее на тормозные области, или чем диффузный свет. Линия (полоска света) эффективна, если она закрывает большую часть центральной области и только малую часть периферии. Поскольку поля этих клеток имеют радиальную симметрию, реакция на такие линии не зависит от их ориентации. Суммируя, можно сказать, что ганглиозные клетки сетчатки и клетки коленчатого тела, т. е. клетки, снабжающие входными сигналами зрительную кору, - это клетки с концентрическими рецептивными полями, центр и периферия которых антагонистичны. Они занимаются в первую очередь не оценкой уровней освещенности, а скорее сравнением уровня освещенности в некоторой небольшой области поля зрения со средней освещенностью ее непосредственного окружения.
Первое из двух основных преобразований, выполняемых зрительной корой, - это такое перераспределение входной информации, в результате которого большинство клеток начинает отвечать не на пятна, а на определенным образом ориентированные отрезки линий. В коре имеется много разнообразных типов клеток; по свойствам своих реакций некоторые из них просты, некоторые сложны, и при знакомстве с ними довольно скоро возникает впечатление, что имеется определенная иерархия, причем более простые клетки подают свои сигналы на вход более сложных. У обезьяны прежде всего выделяется группа клеток, которые ведут себя (насколько это известно) как клетки коленчатого тела: они имеют поля с радиальной симметрией. Все такие клетки располагаются в нижней части одного слоя, называемого слоем IV; он является как раз тем слоем, в который поступает львиная доля волокон коленчатого тела. Создается впечатление, что именно эти наименее сложно устроенные кортикальные клетки являются клетками, ближе всего стоящими ко входу.
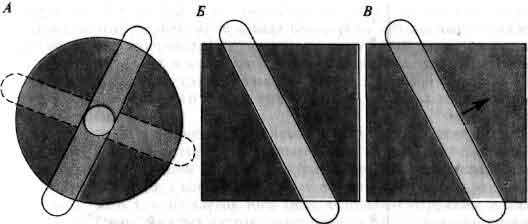
Сравнение рецептивных полей различных клеток зрительного пути. Ганглиозные клетки сетчатки и нейроны латеральных коленчатых тел имеют концентрические поля либо с возбуждающим центром и тормозной периферией (А), либо наоборот. Пятно света, падающее на центр, вызывает реакцию такой клетки; реакцию вызывает и полоса света, падающая на поле в любой ориентации, если она проходит через центр. В зрительной коре существует иерархия нейронов со все усложняющимися характеристиками. Кортикальные клетки, которые получают сигналы прямо из коленчатых тел, имеют поля с радиальной симметрией. Однако кортикальные клетки, расположенные по зрительному пути дальше, реагируют только на линейные стимулы, имеющие определенную ориентацию. Простые клетки (Б) реагируют на такие линейные стимулы лишь в том случае, если они появляются в определенной части поля. Сложные клетки (В) реагируют на определенным образом ориентированные линии независимо от их расположения в поле, и особенно на линии, которые движутся в определенном направлении (стрелка).
За пределами слоя IV все клетки лучше всего реагируют на отрезки линий, ориентированные определенным образом. Типичная клетка реагирует только тогда, когда свет падает на определенную часть поля зрения, но освещение этой части диффузным светом дает малый или нулевой эффект, и небольшие пятна света тоже ненамного лучше. Самая сильная реакция получается, когда нужным образом ориентированная линия вспыхивает в этом месте или, в случае некоторых клеток, поступательно движется через этот участок. Наиболее эффективная ориентация варьирует от клетки к клетке и обычно определена довольно строго, так что отклонение на 10 или 20 градусов по или против часовой стрелки заметно уменьшает реакцию или уничтожает ее. (Остроту этой избирательности представить трудно. Если 10-20° звучит как огромный диапазон, вспомните, что угол на часах между 12 и 1 равен 30°.) Линия, ориентированная под 90° к оптимальному направлению, почти никогда не вызывает реакции.

Позиции рецептивных полей кортикальных нейронов (1-9), картированных при погружении электрода примерно под прямым углом к поверхности, очень близки (А), хотя поля имеют различные размеры и некоторый разброс. При наклонном погружении (5) было зарегистрировано по 2-4 клетки в каждой из четырех точек (1-4) на расстоянии 0,1 мм. В каждой группе попадаются разные размеры и имеется некоторый разброс, но теперь заметен также систематический сдвиг: поля последовательных групп клеток несколько смещены относительно друг друга.
В зависимости от конкретной клетки лучшим стимулом может быть яркая линия на темном фоне или наоборот: это может быть и граница между светлой и темной областями. Если это линия, ее толщина обычно существенна; увеличение ее свыше некоторого оптимального значения уменьшает реакцию точно так же. как увеличение диаметра пятна в случае ганглиозных клеток или клеток коленчатого тела. При этом для данной части поля зрения диаметры центральных зон рецептивных полей клеток коленчатого тела и оптимальные для коры толщины линий сравнимы.
Чувствительные к ориентации нейроны варьируют по своей сложности. Самые простые, которые мы назовем «простыми» клетками, ведут себя так, как будто к ним на вход поступают сигналы непосредственно от нескольких клеток с концентрическими полями, имеющими радиальную симметрию, т. е. от клеток того типа, которые обнаружены в слое IV. Свойства этих простых клеток, реагирующих на оптимально ориентированные линии, предъявляемые в строго определенном месте, легче всего объяснить, приняв, что все центры соответствующих полей входных клеток являются либо возбуждающими, либо тормозными и что они располагаются вдоль прямой линии. В настоящее время у нас нет непосредственных подтверждений этой схемы, но она привлекает своей простотой и тем, что согласуется с рядом косвенных свидетельств. Согласно работе Дж. Ланд (J. Lund) из Медицинской школы Вашингтонского университета, которая за последние несколько лет сделала больше, чем кто-либо другой для изучения этой области коры методом Гольджи, клетки слоя IV проецируются в слои, находящиеся прямо над ними, т. е. примерно туда, где обнаружены простые клетки.
Вторая группа чувствительных к ориентации нейронов - это гораздо более многочисленные «сложные» клетки. Они подразделяются на несколько категорий, но основная их особенность - это то, что местоположение линий для них не так существенно. Сложные клетки ведут себя так, как будто они получают сигналы от некоторого числа простых клеток, имеющих рецептивные поля с одинаковой оптимальной ориентацией, но несколько различающейся локализацией. Эта схема хорошо объясняет сильную стабильную импульсацию, возбуждаемую в сложных клетках, когда линию держат в оптимальной ориентации и проводят через рецептивное поле. При оптимальной ориентации линии многие клетки предпочитают одно из направлений движения другому, противоположному. Для объяснения такого поведения было предложено несколько возможных схем, но точный механизм пока неизвестен.
Хотя нет прямых доказательств того, что чувствительные к ориентации клетки имеют какое-либо отношение к зрительному восприятию, весьма соблазнительно думать, что они представляют некую раннюю ступень анализа зрительных образов. Имеет смысл спросить, от каких клеток этой ранней ступени можно ожидать реакции на какой-нибудь очень простой зрительный стимул, скажем темное пятно на светлом фоне. Любая клетка, рецептивное поле которой окажется полностью внутри или вне границ такого стимула, совершенно не будет затронута его присутствием, поскольку клетки коры эффективно игнорируют изменения диффузной засветки всего рецептивного поля.

Показаны предпочтительные ориентации 23 нейронов, встретившихся при наклонном погружении микроэлектрода в кору (А); самый эффективный наклон стимула неуклонно сдвигался в направлении против часовой стрелки. Б. График сходного эксперимента, однако в этом случае несколько раз наблюдалась инверсия направления сдвига; результаты многих таких экспериментов в сочетании с тем фактом, что микроэлектрод, проникающий в кору под прямым углом, регистрирует только клетки, предпочитающие одну и ту же ориентацию (кроме клеток слоя IV, имеющих поля с радиальной симметрией и не имеющих предпочтительных направлений), заставляют предполагать, что кора подразделена на приблизительно параллельные подслои ткани, причем каждый подслой, называемый ориентационной колонкой, содержит нейроны со сходной ориентационной избирательностью (В).
Единственными клетками, которые должны прореагировать, будут те, через поля которых пройдет граница. Из клеток с радиальной симметрией сильнее всего возбудятся те, у которых граница коснется центра (так как тогда возбуждающая и тормозная зоны будут максимально неравно освещены). Из клеток, чувствительных к ориентации, должны активироваться только те, чьим оптимальным ориентациям случилось совпасть с превалирующим направлением границы. И среди таких клеток простые клетки более требовательны к стимулу, чем сложные, так как они реагируют оптимальным образом только тогда, когда граница пройдет вдоль линии, разделяющей возбуждающую и тормозную зоны. Важно представлять себе, что эта область коры производит свои операции только локально, на маленьких участках изображений; как анализируется или обрабатывается мозгом все изображение, т. е. как эта информация комбинируется и обобщается на более поздних ступенях, если действительно это имеет место, пока неизвестно.
Вторая важная функция зрительной коры обезьяны - комбинировать входы от двух глаз. В латеральном коленчатом теле любой нейрон может реагировать либо на стимуляцию левого глаза, либо на стимуляцию правого, но ни одна клетка не реагирует на стимуляцию обоих глаз. Это может показаться удивительным, поскольку в каждое коленчатое тело поступают сигналы от обоих глаз; однако коленчатые тела устроены таким образом, что пути от двух глаз проходят через них, не смешиваясь. Каждое коленчатое тело подразделяется на шесть слоев, причем три слоя, связанные с левым глазом, переплетаются, как пальцы, с тремя слоями, предназначенными для правого глаза. В каждом слое представлена карта контралатеральной половины зрительного мира (причем все шесть карт располагаются строго упорядоченно, так что на радиальном пути, пересекающем шесть слоев, рецептивные поля всех клеток, которые при этом встретятся, будут иметь практические идентичные позиции в поле зрения). Поскольку каждый данный слой получает входные сигналы только от одного глаза, отдельные клетки каждого слоя должны быть монокулярными.
Даже в зрительной коре нейроны, на которые клетки коленчатого тела переключаются непосредственно - клетки с радиальной симметрией из слоя IV, - все (насколько нам известно) строго монокулярны; то же самое справедливо для всех простых клеток. Только на уровне сложных клеток пути от двух глаз сходятся, но и там смешение информации неполное и принимает специфическую форму. Примерно половина всех сложных клеток монокулярна в том смысле, что любая такая клетка может быть активирована только стимуляцией какого-то одного глаза. На остальные клетки могут оказывать независимые влияния оба глаза.
Если построить карты рецептивных полей бинокулярной клетки для правого и левого глаза (посылая стимулы сначала в один глаз, а затем в другой) и сравнить эти два поля, то окажется, что они имеют одинаковые позиции, уровни сложности, предпочтительные ориентации и направления - т. е. все, что можно узнать о клетке, стимулируя один глаз, подтверждается при стимуляции второго. Есть только одно исключение: если сначала один, а затем второй глаз тестировать идентичными стимулами, две реакции не обязательно будут одинаковыми количественно; во многих случаях один глаз доминирует, устойчиво вызывая более частые разряды, чем второй.
Перебирая клетки, можно найти все степени доминирования глаз, от полной монополии одного глаза через равенство до исключительного влияния второго глаза. У обезьяны клетки с заметным предпочтением одного глаза встречаются несколько чаще, чем клетки, в реакции которых оба глаза вносят примерно равные вклады. По-видимому, связи типичной бинокулярной клетки первичной зрительной коры с двумя глазами практически одинаковы по организации, но могут различаться по числу соединительных волокон.