Тестостерон синтезируется клетками Лейдига, клетками коры надпочечников путем последовательного превращения холестерина липопротеидов низкой плотности в прегненолон, дегидроэпиандростерон и андростендион. Возможно его образование в жировой и других тканях путем взаимопревращения стероидных молекул.
Рисунок 1. Структурная формула тестостерона

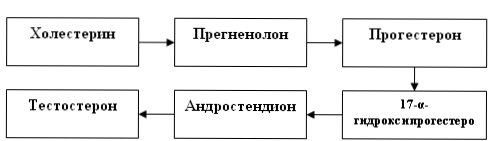
Биосинтез тестостерона
Предшественником тестостерона является холестерол. Источником холестерола служат его эфиры, поступающие в клетку в составе ЛПНП или депонированные в клетке. Превращение холестерола в прегненолон в надпочечниках, яичниках и семенниках происходит идентично, но под действием разных ферментов: АКТГ в надпочечниках и ЛГ в яичниках и семенниках. Освобождение холестерола из его эфиров и синтез кортикостероидов стимулируются кортикотропином. Основными кортикостероидами, обладающими выраженной гормональной активностью, являются кортизол - главный представитель группы глюкокортикоидов, альдостерон - основной минералокортикоид и андрогены. Скорость-лимитирующим этапом является отщепление боковой цепи холестерола.
На первом этапе синтеза кортикостероидов происходит превращение холестерола в прегненолон путем отщепления 6-углеродного фрагмента от боковой цепи холестерола и окисления углеродного атома С20. Прегненолон превращается в прогестерон (С21 предшественник стероидов - кортизола и альдостерона) и стероиды С19 (предшественники андрогенов). Каким именно стероидом окажется конечный продукт, зависит от набора ферментов в клетке и последовательности реакций гидроксилирования.
Превращение прегненолона
в тестостерон эстрон (фолликулин или Е1) протекает с участием пяти ферментов:
1) 3-бета-гидроксистероид-дегидрогеназы (3-бетаОН-СД),
2) дельта-5,4-изомеразы,
3) 17-альфа-гидроксилазы,
4) С17-20 лиазы,
5) 17-бета-гидроксистероид-дегидрогеназы (17-бета-ОН-СД). Соответствующая последовательность реакций получила название прогестеронового пути
Рисунок 2. Синтез тестостерона, путь 1

Рисунок 3. Синтез тестостерона, путь 2
Секретируемый клетками Лейдига (семенники) тестостерон циркулирует в крови в свободной и связанной с белками формах.
Большая часть образованного тестостерона секретируется в семенные протоки, где он соединяется с андроген-связывающим белком (АСЕ), вырабатываемым клетками Сертоли (извитые канальцы семенников). Это депо связанного с белком тестостерона обеспечивает высокую локальную концентрацию андрогена, необходимую для нормального сперматогенеза. Проникнув в клетку органа-мишени, тестостерон трансформируется в свои активные производные, способные обеспечивать на клеточном уровне биологическое действие этого гормона.
ГИПОТАЛАМО-ГИПОФИЗАРНАЯ СИСТЕМА И АНДРОГЕНЫ
В запуске секреции тестостерона яичками участвуют следующие гормоны:
· Гонадотропин-релизинг гормон (ГнРГ);
· Лютеинизирующий гормон (ЛГ);
· Фолликулостимулирующий гормон (ФСГ).

ГнРГ секретируется в гипоталамусе, и ответственен за контроль импульсивной секреции гонадотропинов. Гонадотропины – ЛГ и ФСГ, в свою очередь стимулируют выработку андрогенов и сперматогенез яичками. ЛГ ответственен за регулировку синтеза половых гормонов клетками Лейдига, а ФСГ индуцирует процессы созревания сперматозоидов, а также усиливает стимуляцию клеток Лейдига по отношению к ЛГ, посредствам увеличения ЛГ-рецепторов на мембранах клеток.
Интенсивность выделения гонадотропинов по принципу отрицательной обратной связи зависит от уровня тестостерона и ингибина. Известно, что тестостерон подавляет секрецию ЛГ, а ингибин является ингибитором секреции ФСГ. Это объясняет, почему одним из побочных эффектов заместительной терапии гормональными препаратами у мужчин, а также применение данных препаратов в бодибилдинге, является подавление секреции собственного тестостерона, и при длительном применении вызывает уменьшение яичек в размере, ввиду временного угнетения их функций.
Существует предположение, что в механизме ингибирующего влияния тестостерона на ЛГ участвует не сам тестостерон, а другие стероиды, в которые он трансформируется, например ДГТ и эстрадиол – наиболее активный женский половой гормон. Доказано, что ингибирующее свойство эстрадиола на ЛГ, превосходит ингибирующее свойство тестостерона на ЛГ. Такое положение определяет снижение половых гормонов при ожирении мужчин, связанное с повышенной ароматизацией андрогенов в эстрогены (см. подробнее об ожирении и тестостероне). Ингибирующее свойство эстрогенов по отношению к ФСГ проявляется еще в большей степени, чем к ЛГ.
Для поддержания стероидогенной активности в клетках Лейдига в присутствие ЛГ необходим еще один гормон гипофиза – пролактин. Он ответственен за увеличение число рецепторов ЛГ. Однако это касается только нормального его уровня – при повышенном уровне пролактина угнетается секреторная функция клеток Лейдига. Постоянно повышенный пролактин в плазме крови рассматривается как гиперпролактинемия у мужчин.
Поскольку секреция гонадотропинов имеет импульсивный характер, с наибольшим пиком в утренние часы, то и максимальный уровень тестостерона приходится на 6:00-8:00 часов утра, постепенно снижаясь в течение дня и достигая своего минимума к 20:00-22:00 часам.
Синтез половых гормонов
Стероидные гормоны синтезируются из холестерина (27 углеродных атомов), а холестерин синтезируется из ацетил-КоА.
Холестерин превращается в стероидные гормоны в результате следующих реакций:
- отщепление бокового радикала
- образование дополнительных боковых радикалов в результате реакции гидроксилирования с помощью специальных ферментов монооксигеназ (гидроксилаз) - чаще всего в 11-м, 17-м, и 21-м положениях (иногда в 18-м). На первом этапе синтеза стероидных гормонов сначала образуются предшественники (прегненолон и прогестерон), а затем другие гормоны (кортизол, альдостерон, половые гормоны). Из кортикостероидов могут образоваться альдостерон, минералокортикоиды.