части аппарата Гольджи j
Рис. 3.7. Модель биосинтеза ТТГ, показывающая последовательность
процессов, происходящих в шероховатом эндоплазматическом
ретикулуме, проксимальной и дистальной частях аппарата Гольджи,
образование секреторных гранул и везикул (пузырьков) и продукты
секреции. Показана также секреция тиротрофом интактных а-субъединиц
тиротропина в избыточном количестве. В шероховатом
эндоплазматическом ретикулуме происходит котрансляционное
высвобождение сигнальных пептидов (волнистые линии) и присоединение олигосахаридных остатков к нативным синтезированным de novo полипептидным цепям субъединиц. К р-субъединице присоединяется 1, а к ос-субъединице 2 насыщенных маннозой олигосахаридных радикала. Обычно на свободном конце олигосахаридные цепочки содержат 3 остатка глюкозы. Соединение гликозилированных субъединиц начинается в шероховатом эндоплазматическом ретикулуме. Затем происходит высвобождение молекул маннозы из связи с различными остатками аспарагина в составе субъединиц. В активных тиротрофах в комплексе Гольджи происходит также присоединение остатков фукозы к молекуле тиротропина. В проксимальной части комплекса Гольджи в состав молекулы включаются остатки галактозы, N-ацетилгалактозамина и других углеводов, что приводит к образованию резистентных к действию эндопептидаз форм субъединиц. Здесь же свободные ос-субъединицы подвергаются дополнительному 0-гликозилированию. В дистальной части комплекса Гольджи присоединяются остатки серной и/или сиаловой кислот. Наконец, гетеродимерная молекула ТТГ секретируется путем экзоцитоза. (Werner and Ingbar's The Thyroid, 1996)
Регуляция функции щитовидной железы

|
Y I
т Y_
I
X?
Y
чх?
т
| к! |
ЧХ? /
I 4
• 1 л і »*
Ні ЇЇЇ
| ■Р- | mm 1! | ||
| Н | |||
| X? | V | ||
| о-в | X | ||
| t | |||
| Ах | АН | ||
| / | И | .Ы. Ті 1 1 | |
| ? т | |||
| ** | V | ||
| і | |||
| т | А» | ||
| "ч | * | ■ * | |
| h | t | ||
| і/ | V | ||
| - І | |||
| * |
Jti
JtL
itL
Рис. 3.8. Схематическое представление последовательности присоединения остатков серной (сульфатирование) и сиаловых кислот к олигосахаридным цепочкам, связанным с аспарагином в составе молекулы тиротропина. Олигосахаридные цепочки содержат большое количество остатков маннозы и 3-х молекул глюкозы. Интактная молекула активируется при удалении сначала двух гликозидных остатков, а затем замещения 3-го N-ацетилглюкозамином. Ключевой точкой в разветвлении цепи является ацетилирование/замещение остатков глюкозы, которое приводит к образованию N-ацетилгалактозамина (GalNAc). Остатки галактозы затем могут присоединять остатки сиаловых кислот, a GalNAc -подвергаться сульфатированию. S-1, S-2 и S-3 обозначает присоединенное количество сульфатов, N-1, N-2 и N-3 - остатков сиаловых кислот; S-N обозначает олигосахарид, одна цепочка которого сульфатирована, а другая содержит остаток сиаловой кислоты. (Baenziger JU, Green ED, Biochem Biophys Acta, 1988).
Известны случаи наличия у детей точечных мутаций гена,
кодирующего синтез (3-цепи ТТГ. Следствием такого аутосомного
рецессивного нарушения является невозможность структурно
измененной (3-субъединицы вступать во взаимодействие с ос-субъединицей и образовывать биологически активный тиротропин. У детей с подобной патологией наблюдаются клинические признаки гипотиреоидизма.
Концентрация ТТГ в крови колеблется с периодом около 1,8 часа и достигает своего максимума в интервале между полуночью и 4-мя часами утра. Нормальные пределы колебаний содержания ТТГ в крови составляют от 0,5 до 5,0 мкЕд/мл. Секреция ТТГ минимальна в послеобеденные часы суток. Большая разница в концентрациях ТТГ в
Регуляция функции щитовиднойжелезы
различное время суток не оказывает существенного влияния на концентрации Т4 и Т3 в циркуляторном русле, поскольку в организме имеется большой пул внетиреоидного Т4. Время полужизни ТТГ в плазме крови составляет около получаса, а его продукция за день равна около 40-150 мЕд. Уровень ТТГ повышается при первичном гипотиреоидизме, может возрастать после частичной тиреоидэктомии и (или) после радиойодтерапии новообразований щитовидной железы. Он понижается при гипертиреоидизме, может быть пониженным или нормальным при вторичном гипотиреоидизме. Кроме ТТГ в сыворотке крови может также содержаться а-субъединица тиротропина в количестве около 0,5-2,0 мкг/л. Ее концентрация может быть более высокой при ТТГ-секретирующих опухолях гипофиза и у женщин после наступления менопаузы.
Синтез ТТГ осуществляется в тиротрофах переднего отдела гипофиза. Эти клетки составляют около 5% от других клеточных популяций. Синтез тиротропина регулируется многими биологически активными веществами (табл. 3.2). Среди них особо важное место занимают тиреоидные гормоны и ТРГ гипоталамуса (действие ТРГ описано в предыдущем разделе).
Согласно классической схеме, тиреоидные гормоны по каналу отрицательной обратной связи подавляют синтез тиротропина, действуя как непосредственно на сами тиротрофы, так и через уменьшение секреции ТРГ гипоталамусом. Синтез и секреция ТТГ подавляется при повышении уровня Т4 и Т3 в сыворотке крови (например, при гипертиреоидизме) и стимулируется при понижении уровня тиреоидных гормонов (например, при гипотиреоидизме). При повышении содержания Т3 внутри тиротрофа резко уменьшается уровень мРНК как |3-, так и а-субъединиц, но более чувствительной к этим изменениям является мРНК Р-субъединицы. Так, при длительном действии Т3 содержание мРНК этой субъединицы в тиротрофах уменьшается до следовых количеств. Этот эффект достигается тиреоидными гормонами на уровне торможения скорости транскрипции гена, которая падает после связывания гормонов с их рецепторами.
Через изменение числа рецепторов может регулироваться общая и пороговая чувствительность тиротрофов к действию как ТРГ, так и тиреоидных гормонов. Таким образом, их действие на синтез и секрецию ТТГ является взаимодополняющим и коррегирующим.
Биогенные амины серотонин и дофамин могут ингибировать секрецию ТТГ гипофизом. Введение антител к серотонину увеличивает базальный уровень ТТГ и потенцирует ответную реакцию повышения уровня ТТГ на действие холода. Базальный уровень секреции ТТГ, равно как и уровень секреции, стимулируемый действием ТРГ, быстро уменьшается под влиянием дофамина и его агониста бромокриптина. Это является результатом быстрого (в течение нескольких десятков минут) торможения дофамином транскрипции генов обоих субъединиц ТТГ. Дофамин уменьшает внутриклеточный уровень цАМФ, от которого также зависит экспрессия генов, кодирующих синтез ТТГ.
Табл. 3.2. Факторы, контролирующие синтез и секрецию гормонов тиреоидной системы
| Гипоталамус: синтез и секреция ТРГ | Передний гипофиз: синтез и | Щитовидная железа: синтез и | ||||
| секреция | ТРГ | секреция тиреоидных гормонов | ||||
| Стимулируют | Ингибируют | Стимулируют | Ингибируют | Стимулируют | Ингибируют | |
| • Понижение | • Повышение | • ТРГ | • | Повышение | • ТТГ | • Антитела, |
| уровней Т4 и Т3 | уровней Т4 и Т3 | • Понижение | уровней | • Антитела, | блокиру- | |
| в сыворотке | в сыворотке | уровней | Т4иТ3 | стимули- | ющие | |
| и Тз в нейронах | и Тз в нейронах | Т4иТ3 | в сыворотке | рующие | рецептор | |
| • Действие | • а-адрено- | в сыворотке | и Тз в | рецептор | ТТГ | |
| холода | блокаторы | и Тз в | тиротрофах | ТТГ | • Избыток | |
| (новорожденные | • Опухоли | тиротрофах | • | Повышение | йода | |
| и животные) | гипоталамуса | Снижение | активности | • Литий | ||
| • Катехоламины | активности | дейодиназы | ||||
| (а-адре но- | дейоди- | 5'-ДП | ||||
| миметики) | назы 5'-Дії | • | Соматостатин | |||
| • Аргинин- | • Эстрогены: | • | Дофамин, | |||
| вазопрессин | увеличивают плотность ТРГ связывающих мест | • • • | агонисты рецепторов дофамина (бромо- криптин) Глюкокор- тикоиды Хронические заболевания Опухоли гипофиза |
Регуляция функции щитовидной железы
Подобным, ингибирующим действием на базальную секрецию ТТГ и на секрецию, стимулируемую действием ТРГ, обладает также и соматостатин. Его ингибирующее действие более выражено при низких концентрациях тиреоидных гормонов.
Соматостатин может связываться в гипофизе и других структурах мозга с несколькими типами специфических, высокоаффинных к нему рецепторов. При этом свое ингибирующее действие на секрецию ТТГ он может оказывать либо через систему циклических мононуклеотидов, либо вызывая гиперполяризацию мембраны тироцитов через изменение состояния потенциалзависимых калиевых каналов, что в конечном итоге ведет к снижению внутриклеточного уровня кальция.
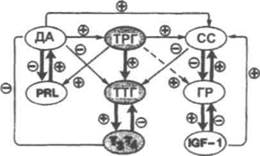
Рис. 3.9. Схема комплекса взаимодействий между первичными и вторичными модуляторами функции тиротрофов. ДА - дофамин; СС-соматостатин; ГР - гормон роста; PRL - пролактин; IGF-1 -инсулиноподобный фактор роста-1; «+» стимуляция, «-» угнетение. (Werner and Ingbar's «The Thyroid», 1996)
Цитокины интерлейкин-1 (ИЛ-1) и в меньшей степени интерлейкин-6 (ИЛ-6) вызывают преходящее уменьшение содержания в плазме крови ТТГ и общих уровней Т4 и Т3, но увеличивают уровень свободного Т4 Снижение в крови общего уровня тиреоидных гормонов является, как полагают, результатом прямого ингибирующего влияния этих цитокинов на функцию тиротрофов и ингибирующего действия на образование гормонов щитовидной железой. Кроме того, ингибирующее действие ИЛ-1 может быть также результатом его влияния на секрецию в гипоталамусе соматостатина или ТРГ.
Ингибирующим эффектом на секрецию ТТГ обладают норадреналин и глюкокортикостероиды, что может иметь отношение к регуляции уровня ТТГ и функции щитовидной железы при стрессе, а также к суточному ритму изменения уровня ТТГ. Острые или хронические заболевания также могут сопровождаться подавлением секреции ТТГ с последующим восстановлением при выздоровлении. Существование зависимости образования и секреции ТТГ от действия некоторых лекарственных средств, состояния стресса и наличия у человека соматических заболеваний должно учитываться врачом при интерпретации результатов исследования уровня тиротропина.
Регуляция функции щитовидной железы
SS 0A2 odAD ТРГ