Бесполое размножение реализуется путём образования конидий и спорангиоспор, содержащих весь генетический материал, необходимый для возникновения и развития новой колонии. Если у гриба неизвестны половые формы, то строение конидий используют как классифицирующий признак. Грибы, не размножающиеся половым путём, или виды, для которых совершенные (половые) формы неизвестны, объединены в класс Deuteromycetes, или Fungi imperfecti (несовершенные грибы).
Конидии — неполовые репродуктивные структуры. Они могут быть одноклеточными (микроконидии) либо многоклеточными (макроконидии). У некоторых грибов конидии формируются на концах конидионосных гиф— конидиофоров (бластоконидии), у других— по бокам и на концах обычных гиф (таллоконидии) путём их септирования и расчленения. Бластоконидии (бластоспоры) формируются в результате отпочковывания от конидиофора. Их образование характерно для дрожжей и дрожжеподобных грибов. Наиболее типичные таллоконидии — артро- и хламидоконидии.
Артроконидии (артроспоры) образуются в результате фрагментации септированных гиф, распадающихся на отдельные клетки (рис. 2-10). Хламидоконидии (хламидоспоры) — таллоконидии, образующиеся внутри нитей мицелия -в виде увеличенных клеток, окружённых толстой оболочкой. За счёт плотной оболочки хламидоконидии приобретают устойчивость к различным воздействиям и сохраняются после гибели и распада мицелия (отдалённо напоминая этим цисты простейших). Выделяют хламидоконидии терминальные (концевые), располагающиеся на конце гиф, и интеркалатные (промежуточные), располагающиеся в середине гифы.
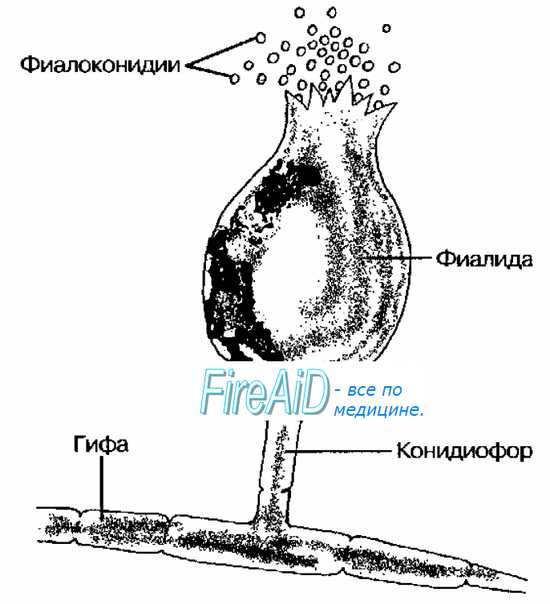
Рис. 2-11. Бесполое размножение грибов с образованием фиалоконидии.
Спорангиоспоры — споры, образующиеся вегетативным путём в специализированных мешках (спорангиях). Последние располагаются на концах особых гиф — спорангиофорах. К основным типам спорангиев относят фиалиды {образования, напоминающие бокал), содержащие фиалоспоры, или фиалоконидии (рис. 2-11) и аннелиды (кольцевидные структуры), содержащие аннелоспоры, или аннелоконидии.
Бесполое спороношение грибов. Наиболее примитивным органом бесполого размножения у низших грибов является зооспорангий, представляющий собой расширенное окончание гифы, внутри которого формируются подвижные споры с одним или двумя жгутиками – зооспоры.Более совершенной формой бесполого размножения низших грибов является образование спорангиев – шаровидных вместилищ, на концах ответвлений грибницы. Ветвь, несущая спорангий, называется сиорангиеносцем. Внутри спорангия формируются неподвижные споры – спорангиоспоры. Самой распространенной формой бесполого размножения, свойственной высшим грибам, является конидиальное спороношение. Конидии – это споры, образующиеся на концах вегетативных гиф или на конечных ответвлениях специальных органов – конидиеносцев.Конидиеносцы и конидии очень разнообразны по форме, размерам, строению и окраске, а также по характеру развития и размещения.Бесполое спороношение у фитопатогенных грибов обычно образуется многократно в течение вегетационного периода и служит для массового распространения гриба и повторного заражения растений.
Половое размножение грибов
Среди грибов, патогенных для человека, способность размножаться половым путём выявлена у представителей классов Ascomycetes, Basi-diomycetes и Zygomycetes. Половым путём образуются аскоспоры, базидиоспоры и зигоспоры.Аскоспоры. У аскомицетов половым путём в специализированных булавовидных (реже шаровидных) клетках — асках — образуются споры, число которых всегда кратно двум.

Базидиоспоры образуются после мейотического деления спор на базидии — концевой клетке мицелия. Обычно на базидии развивается четыре споры. Зигоспоры. Зигомицеты могут размножаться как вегетативным, так и половым путём. При половом размножении верхушки близко расположенных гиф соединяются, их оболочки растворяются, ядра сливаются (как и у прочих эукариотов). Процесс проходит постадийно: первоначально сливаются два протопласта (плазмогамия), затем оба ядра сопряжённо делятся и после образования плодового тела сливаются (кариогамия). За кариогамией следует мейоз — редукционное деление, при котором число хромосом уменьшается до исходного гаплоидного. Затем образуется зигоспора с толстыми ригидными стенками. Процесс образования зигоспор известен как зигогамия.

половое размножение у грибов бывает различных типов. Сущность его заключается в том, что происходит слияние двух половых клеток (гамет) - мужской и женской - или двух вегетативных талломов, функционирующих как половые клетки, в результате возникает новообразование (зигота). Сливающиеся гаметы содержат только половинный набор хромосом. В зиготе число хромосом соответственно удваивается. Гаметы являются структурами, которые находятся, имея половинный хромосом, в гаплоидной фазе, а зигота переходит уже в диплоидную фазу.У высших грибов половой процесс протекает как слияние органов и клеток, не дифференцированных на гаметы. Образовавшаяся в результате слияния зигота (также не дифференцированная и обычно представляющая собой лишь соответствующее ядерное состояние) без периода покоя переходит к дальнейшему развитию; в ней формируются дикарионы ядер противоположных полов, которые потом попарно сливаются и претерпевают редукционное деление. Гаплоидные ядра, которые образовались в процессе редукционного деления, переходят в аскоспоры, образующиеся в сумках или в базидиоспоры, образующиеся на специальных клетках - базидиях - базидиомицетах экзогенно.
16. Споры грибов, развивающиеся в период спороношения, могут отличаться. Одни споры - пропагативные - развиваются в огромном количестве, служат для быстрого расселения грибов. Так, в плодовом теле дождевика - до 7,5 миллионов спор, а шампиньон за 5 дней рассеивает более 10 биллионов спор. Такие споры быстро прорастают, но являются маложивучими, часто недозрелыми и в больших количествах погибают. Другие споры - покоящиеся - развиваются в малом количестве, нуждаются в покое, во время которого происходит их дозревание, служат для сохранения вида при неблагоприятных условиях. Такие споры могут сохраняться до 10-12 лет и прорастают медленно. Прорастают они обычно спорангием, дающим массу спор.
17. Царство грибов характеризуется разнообразием жизненных циклов и вариантов ядерного статуса (плоидность, количество ядер в клетке, их генетическая разнородность или однородность). У различных таксономических групп грибов выделяют до 7 типов жизненного цикла (по Дж. Рэйперу [Raper]).
1. Бесполый цикл характерен для нескольких десятков тысяч видов аскомицетов ибазидиомицетов, утративших половую стадию — так называемых дейтеромицетов.
2. Гаплоидный цикл известен у зигомицетов, многих хитридиомицетов. Мицелий содержит множество гаплоидных ядер (мультикариотический мицелий), которые делятся митотически по мере роста гиф.
3. Гаплоидный цикл с ограниченным дикарионом характерен для большинства аскомицетов, мицелий их также чаще всего бывает гаплоидный мультикариотический. Гаметы или гаметангии вначале сливаются цитоплазмами (происходит плазмогамия) без слияния ядер (кариогамии) и прорастают дикариотическими гифами, называемыми также аскогенными.
4. Гаплоидно-дикариотический (гапло-дикариотический) цикл встречается у многих базидиомицетов — гименомицетов,гастеромицетов, ржавчинных грибов. Он сходен с предыдущим, но характеризуется длительной стадией дикариона, которая чаще всего бывает доминирующей.
5. Дикариотический цикл характерен для ограниченной группы базидиомицетов — головнёвых грибов. Гаплоидная фаза у них представлена базидиоспорами и прорастающими из них одноядерными споридиями, которые способны расти на питательной среде. Сливаясь попарно, споридии формируют дикариотический мицелий.
6. Гаплоидно-диплоидный (гапло-диплоидный) цикл встречается у низших водных грибов — бластокладиевых, хитридиомицетов. Диплоидный спорофит образует зооспоры, прорастающие в такие же диплоидные спорофиты (бесполое размножение) и мейоспорангии, дающие начало гаплоидному половому поколению — гаметофитам..
7. Диплоидный цикл известен у дрожжей Saccharomyces cerevisiae и других сахаромицетов, кроме настоящих грибов характерен также для оомицетов, например, Phytophthora infestans. Преобладает диплоидная стадия почкующихся клеток, ядра которых в определённых условиях (например, истощение питательной среды) делятся мейозом и формируют гаплоидные аскоспоры..
Часто один и тот же гриб образует в цикле своего развития не одно, а несколько спороношений. Они чередуются, следуя одно за другим, и каждый раз гриб меняет форму. У некоторых грибов и ветегативное тело может видоизменяться на протяжении жизни, переходя в зависимости от условий внешней среды от мицелиального роста к дрожжеподобному и наоборот. Такие изменения формы грибов в течение жизни получили название полиморфизма. Полиморфизм, связанный со сменой дрожжевой и мицелиальной форм роста в ходе вегетативного развития гриба обычно называют диморфизмом, а со сменой разных типов спороношений - плеоморфизмом. Явление плеоморфизма грибов вызывает большие затруднения в работе микологов. Действительно, если не исследовать связи между отдельными спороношениями, каждое из них можно принять за самостоятельный гриб
ПЛЕОМОРФИЗМ— своеобразный тип полиморфизма, проявляющийся в способности одного вида гриба образовывать разные спороношения на различных стадиях своего развития. В жизненном цикле развития этих грибов наблюдается последовательная смена таких спороношений, которые в большинстве случаев резко отличаются друг от друга по морфологии. П. известен у сумчатых, ржавчинных и некоторых др. групп грибов.
18. КЛАСС СУМЧАТЫЕ ГРИБЫИЛИ АСКОМИЦЕТЫ(ASCOMYCETES)
Сумчатые грибы, или аскомицеты,— один из крупнейших классов грибов. В нем более 30 000 видов, что составляет около 30% всех известных видов грибов. Входящие в этот класс грибы чрезвычайно разнообразны по строению. Сюда относятся, например, дрожжи, представленные одиночными почкующимися клетками, и виды с плодовыми телами различной формы и размеров, от микроскопических до крупных, достигающих иногда 10—20 см (сморчки, строчки).
У низших аскомицетов сумки образуются непосредственно на мицелии, а у высших — в специальных вместилищах — плодовых телах (рис. 69). Различают следующие типы плодовых тел: клейстотеций (полностью замкнутое плодовое тело), перитеций (полузамкнутое, обычно кувшиновидпое плодовое тело с отверстием на вершине) и апотеций (открытое, обычно чашевидное плодовое тело, на верхней стороне которого расположен слой сумок и парафиз). Клейстотеций, перитеции и апотеции — настоящие плодовые тела, развитие которых происходит по аскогимениальному типу — образование их оболочки (перидия) происходит одновременно с развитием аскогенных гиф и сумок.Настоящие плодовые тела могут развиваться как непосредственно на мицелии, так и на стромах — плотных сплетениях гиф различной формы, размера и консистенции. Однако они всегда имеют собственный перидий, заметный хотя бы на ранних стадиях развития плодового тела. 
19. Плодовые тела базидиомицетов различны по форме и консистенции. Они могут быть паутинистыми, рыхлыми, плотно-войлочными, кожистыми, деревянистыми, мягкомясистыми, могут иметь форму пленок, корочек, могут быть копытообразными или состоять из шляпки и ножки.
Спороносный слой плодового тела — гимений — располагается у более примитивных видов на верхней стороне плодовых тел, а у более высокоорганизованных — на нижней. Гимений базидиальных грибов состоит из базидий с базидиоспорами и парафиз. У некоторых видов в гимении находятся цистиды — крупные клетки, возвышающиеся над гимениальным слоем. Они защищают гимениальный слой и особенно базидий от давления сверху.
Поверхность плодового тела, несущую гимении, называют гименофором. У низших представителей он гладкий, а у более высокоорганизованных имеет форму зубцов, трубочек, пластинок (рис. 155, 156, 157).

ХИТРИДИОМИЦЕТЫ
(Chytridiomycetes), класс настоящих грибов. Гл. обр. одноклеточные и микроскопич. формы. Вегетативное тело мн. X. представлено плазмодием или зачаточным мицелием (ризомицелий). В отличие от оомицетов клеточные стенки X. не имеют целлюлозы, а в качестве скелетного вещества содержат хитин. Размножение бесполое, при к-ром образуются зооспорангии с одножгутиковыми зооспорами, и половое (гаметогамия, оогамия, соматогамия). Возникающая при половом процессе зигота превращается в покоящуюся спору, прорастающую впоследствии в диплоидный таллом. На талломе после редукционного деления формируются зооспорангии с гаплоидными зооспорами. 3 порядка: хитридиевые (Chytridiales), бластокладиевые (Blastocladiales) и моноблефаридовые (Monoblepharidales), различающиеся между собой вегетативной фазой, микроструктурой жгутиков и характером полового размножения. Св. 90 родов, ок. 500 видов. X. в осн. водные формы, паразитирующие на пресноводных и мор. водорослях, водных грибах, простейших, редко на высших растениях. Немногие — сапротрофы. Нек-рые виды X. (напр., ольпидиум, синхитриум) вызывают болезни с.-х. растений. Хитридиомицеты тесно связаны с водной средой (морской и пресноводной), гдепаразитируют на водорослях и беспозвоночных. Могут вызывать массовую гибель водных организмов вплоть до амфибий. Могут развиваться во влажных почвах и вызывать болезни высших растений: чёрную ножку капусты (Olpidium brassicae), рак картофеля (Synchytrium endobioticum). ругой род хитридиевых грибов, имеющий особо важное практическое значение,— сипхитриум (Synchytrium). Основное его отличие от предыдущего рода заключается в том, что в цикле развития видов этого рода вместо одного зооспорангии развивается от пяти до девяти, скученных вместе (сорус).
Круг хозяев синхитриума очень широк, но все они относятся к высшим наземным растениям. В большинстве случаев при поражении синхитриумом на листьях, черешках, стеблях пораженных растений развиваются небольшие черные или коричневые бородавочки — галлы, представляющие собой вздутия клеток эпидермиса, внутри которых находится паразит. ругой род хитридиевых грибов, имеющий особо важное практическое значение,— сипхитриум (Synchytrium). Основное его отличие от предыдущего рода заключается в том, что в цикле развития видов этого рода вместо одного зооспорангии развивается от пяти до девяти, скученных вместе (сорус).
Круг хозяев синхитриума очень широк, но все они относятся к высшим наземным растениям. В большинстве случаев при поражении синхитриумом на листьях, черешках, стеблях пораженных растений развиваются небольшие черные или коричневые бородавочки — галлы, представляющие собой вздутия клеток эпидермиса, внутри которых находится паразит.
21. Оомицеты (лат. Oomycota, Oomycetes) — группа мицелиальных организмов, включающая 70 родов и 570 видов. Другие являются паразитами высших растений, в частности, к оомицетам относится Phytophthora infestans, вызывающая фитофтороз картофеля. Оомицеты размножаются как бесполым, так и половым путём. При бесполом размножении образуют зооспоры вспорангиях. В ряде случаев спорангий может отделяться от спорангиеносца и прорастать как одна спора. Зооспоры имеют два жгутика — передний перистый и бичевидный задний, собственно и служащий для передвижения.
Половое размножение представлено своеобразной оогамией. Оогоний содержит одну или несколько яйцеклеток, в то время как антеридий не образует обособленных гамет и приоплодотворении просто переливает часть своего клеточного содержимого с ядрами в оогоний, что является характерным признаком, по которому выделена группа. Образующиеся после оплодотворения ооспоры являются покоящейся формой оомицета, в которой он пережидает неблагоприятный период, например, зиму. Весной они прорастают, давая зооспорангий. Таким образом, ооспора к тому же и единственная диплоидная стадия в цикле развития оомицетов. Для оомицетов характерен гинадромиксис (от «гина» — женский, «андрос» — мужской), т.е. смена пола при наличии определенных феромонов (антеридиол, оогониол). Встречается только у двудомных, у которых антеридии и архегонии развиваются на разных талломах. Классификационное положение
Несмотря на наличие развитого мицелия, оомицеты были исключены из числа грибов. Из всех грибов лишь они и хитридиомицеты имеют подвижные зооспоры, оомицеты не имеют хитина в клеточной стенке, биосинтез лизина происходит у них как у растений, а не как у остальных грибов, наконец, они имеют совершенно не свойственную грибам ультраструктуру митохондрий. На основании этих признаков в конце XX векацарство грибов было разделено на два отдела: Oomycota и Eumycota, в наше время отдел Oomycota с классом Oomycetes перенесён в царство Chromista; по другой классификационной схеме класс Oomycetes включается в тип Heterokontophyta царства Protista.
Из группы пероноспоровых в Ленинградской области особенно распространена фитофтора инфестанс, вызывающая фитофтороз картофеля. Гриб поражает и листья, и клубни картофеля. На зараженных листьях образуются крупные бурого цвета пятна, развивающиеся от края листовой пластинки к центру. На границе между больной и здоровой тканью хорошо заметен с нижней стороны листа нежный белый налет. Он состоит из выступающих из устьиц спорангиеносцев, отчленяющих лимоновидные зооспорангии, которые подхватывает ветер и переносит на новые здоровые растения. Кроме картофеля гриб поражает томаты, развиваясь как на листьях, так и на плодах. известной под названием ложной мучнистой росы или пероноспороза. возбудители ложной мучнистой росы капусты (пероноспора капустная), клевера (пероноспора клеверная), лука (пероноспора Шлейдена) и многие другие. Наиболее опасным является пероноспороз капусты, гороха и клевера. Пероноспороз капусты, например, проявляется в течение всей вегетации, но особенно он опасен при поражении рассады и семенников. Пероноспороз гороха и клевера в периоды массовых вспышек заболевания (эпифитотии) может нанести большой ущерб как молодым растениям, так и семенникам. В последнем случае семян чаще всего не образуется, а если они и образуются, то имеют небольшой процент всхожести. На поверхности пораженных органов развиваются конидии. С их помощью гриб распространяется и, попадая на листья нового растения, заражает его.
22. Пероноспоровые по сравнению с сапролегниевыми и лептомитовыми представляют собой следующий этап приспособления грибов к жизни на суше. Это довольно древняя группа, возникновение которой относят к палеозойской эре (карбон), наиболее эволюционировавшая среди ооцицетов. В ней насчитывается несколько сотен видов, причем наибольшее их число входит в семейство пероноспоровых (Peronosporaceae), представители которого характеризуются более совершенными признаками, тогда как другие семейства содержат значительно меньше видов и эволюционно менее прогрессировали.Порядок пероноспоровых делят на 3 семейства: питиевые, цистоповые и пероноспоровые.По приуроченности к среде Обитания виды пероноспоровых грибов можно отнести к гидрофилам, зоофилам и фитофилам.По характеру развития пероноспоровые грибы — эфемеры. Они требуют короткого времени для образования спороношения и затем относительно быстро отмирают, но при помощи своих спор, попавших на благоприятные субстраты, развиваются заново, возрождаясь в новых поколениях, формирующих новые генерации спор, многократно повторяющиеся в течение вегетационного периода. Наибольшего развития пероноспоровые грибы достигают в ранневесенний, весенний и весеннелетний периоды. В это время года, по существу, во всех зонах обитания нарастает численность видов и частота их появления. Некоторый подъем развития пероноспоровых грибов происходит и осенью, что связано с выпадением обильных осадков и смягчением температурного режима.ОСНОВНЫЕ БОЛЕЗНИ ДАННОГО ТИПА ЭТО- Пероноспора табака, Пероноспора шпината,ПРЕНОСПОРА ХМЕЛЯ,КАРТОФЕЛЯ И ТД.