- Содержит более 500 видов. За немногим исключением, почти все представители класса ведут наземный образ жизни. Среди них имеются как сапротрофы, так и паразиты грибов, высших растений, насекомых, других животных и человека.
- Их неклеточный многоядерный мицелий хорошо развит. У некоторых в зрелом состоянии он разделяется на отдельные клетки или с самого начала многоклеточный. В клеточных стенках мицелия содержатся хитин и хитозан.
- Бесполое размножение осуществляется неподвижными спорами одетыми оболочкой или развивающимися внутри особых вместилищ - спорангиев (спорангиоспоры), либо экзогенно на конидиеносцах. В ряде случаев можно проследить эволюцию, связанную с наземным существованием, от размножения при помощи спорангиоспор к размножению при помощи конидий.
- Основная особенность зигомицетов, отраженная в названии, заключается в своеобразном половом процессе - зиготами, при котором сливается содержимое двух клеток (отделяющихся при этом перегородками от несущих гиф), не дифференцированных на гаметы. У некоторых видов эти клетки принадлежат одному и тому же мицелию, у большинства - к разным мицелиям. Явление гетероталлизма было впервые обнаружено, описано и изучено А. Блексли (1904 г.) как раз у представителей зигомицетов.
- Копулирующие клетки большей частью не различимы по величине и внешнему виду, у некоторых же одна из клеток больше, а другая - меньше или одна из гиф несет придатки за пределами перегородки, отделяющей эту клетку, у других этих придатков нет. На месте слияния клеток, функционирующих как гаметангии (так как они многоядерные) или как гаметы (в случае их одноядерности), развивается покоящаяся зигоспора. Иногда такие покоящиеся образования развиваются без слияния клеток (азигоспоры). При прорастании покоящихся спор образуется гифа со спорангием на конце.
- Происхождение зигомицетов возможно от каких-то предков, общих с хитридиевыми грибами, или от потерявших подвижность жгутиковых. С другой стороны, их можно связать с сумчатыми грибами.
- Класс делится на следующие порядки: мукоровые (Mucorales), энтомофторовые (Entomophtorales), эндогеновые (Endogonales), зоопаговые (Zoopagales).
представители мукоровых грибов, напри- мер спородиния крупная, изопус черный И ТД.
24. Сумчатые грибы, или аскомицеты, — один из крупнейших классов грибов. В нем более 30 000 видов, что составляет около 30% всех известных видов грибов. Входящие в этот класс грибы чрезвычайно разнообразны по строению. Сюда относятся, например, дрожжи, представленные одиночными почкующимися клетками, и виды с плодовыми телами различной формы и размеров, от микроскопических до крупных, достигающих иногда 10-20 см (сморчки, строчки). Но все эти многообразные формы связаны общим происхождением и имеют ряд общих черт, на основании которых они объединяются в этот класс.Основной признак аскомицетов — образование в результате полового процесса сумок (или асков) — одноклеточных структур, содержащих фиксированное число аскоспор, обычно 8 (рис. 50). Сумки образуются или непосредственно из зиготы (у низших аскомицетов), или на развивающихся из зиготы аскогенных гифах. В сумке происходит слияние ядер зиготы, а затем мейотическое деление диплоидного ядра и образование гаплоидных аскоспор. У высших аскомицетов сумка представляет не только место образования аскоспор, но и активно участвует в их распространении.Вегетативное тело аскомицетов — разветвленный гаплоидный мицелий, состоящий из многоядерных или одноядерных клеток. В отличие от зигомицетов перегородки (септы) в мицелии аскомицетов образуются упорядоченно, синхронно с делением ядер. Развитие септ происходит от стенок гифы к центру, напоминая сужение диафрагмы в объективе фотоаппарата. В центре септы остается пора (рис. 51), через которую происходит движение цитоплазмы с со скоростью от 1-2 до 25-40 см/ч. Некоторые органеллы клетки, даже ядра, могут мигрировать через поры. Наличие пор в септах играет существенную роль в переносе питательных веществ по гифам в зону роста.
В цикле развития аскомицетов большую роль играет бесполое размножение. Споры бесполого размножения (конидии) образуются на гаплоидном мицелии экзогенно (реже эндогенно) на конидиеносцах различного строения. Конидиальные спороношения аскомицетов очень разнообразны по морфологии. Конидиеносцы образуются одиночно на мицелии, соединяются в пучки (коремии) или подушечки (спородохии), развиваются плотным слоем на поверхности сплетения гиф (ложа) или внутри шаровидных либо грушевидных споровместилищ с отверстием на вершине (пикниды). Типы конидиальных спороношений и различные способы образования конидий подробно онисапы в главе о дейтеромицетах, или несовершенных грибах. Многие представители этого класса имеют большое экономическое значение как продуценты антибиотиков, алкалоидов, ростовых веществ (гиббереллинов), витаминов (рибофлавина и др.), ферментов, кормового белка, а также как возбудители спиртового брожения. Наконец, многие аскомицеты широко используются сейчас в качестве объектов генетических и биохимических исследований.
25. голосумчатые, первичносумчатые (Hemiascomycetidae), подкласс сумчатых грибов. Для Г. характерно развитие сумок непосредственно на мицелии или на почкующихся клетках без образования плодового тела и большей частью изолированно одна от другой. Одни исследователи рассматривают Г. как примитивные организмы, другие — как регрессирующие. По способу образования сумок Г. делят на 2 порядка: Endomycetales и Taphrinales. У Endomycetales сумки развиваются из зиготы, образующейся партеногенетически (см. Партеногенез) или путём копуляции двух грибных клеток. Многие виды этих грибов (дрожжи) вызывают брожение и широко используются в пищевой промышленности. У Taphrinales сумки образуются из двуядерных аскогенных клеток, развивающихся, подобно хламидоспорам, из двуядерного мицелия. Это паразиты растений, вызывающие болезни: курчавость листьев персика, миндаля, вишни, груши, берёзы, ольхи, тополя, "кармашки" сливы и алычи, "ведьмины метлы" сливы, алычи, берёзы, вздутие листьев лещины. Голосумчатые грибы (Гемиаскомицеты). Немногочисленный подкласс. К нему относятся примитивные аскомицеты, у которых отсутствуют плодовые тела и сумки развиваются одиночно или слоем непосредственно на мицелии. Включает 3 порядка: эндомицетовые (как правило, отсутствует мицелий, клетки размножаются почкованием, редко делением), тафриновые (обдадают эутуникатной сумкой, с открывающейся крышечкой, которая образуется не влодовых телах - они утрачены в результате приспособления к паразитизму, - а на мицелии, плотным слоем под кутикулой растения-хозяина; в цикле развития преобладает дикарионтическая фаза), протомицетовые (группа представляет боковую слепую эволюционную ветвь).
26. ПЛЕКТОМИЦЕТЫ
— группа примитивных аскомицетов, имеющих ангиокарпные плодоношения без остиоли, вся полость которых выполнена аскогенными гифами, которые дают шаровидные аски, без парафиз и др. нитей. Аски неправильно разбросаны в псевдопаренхиматозной ткани. Группа объединяет сумчатые грибы пор. Eurotiales, Microascales, Onygenales.
27. Порядок Эризифовые или Мучнисторосяные грибы (Erysiphales)
Своеобразная группа грибов-паразитов. Большинство их живет на поверхности различных органов растений, где развивается их белая, часто затем темнеющая, грибница со спороношениями. Питание мучнисторосяных грибов осуществляется при помощи гаусториев, проникающих в ткани растений. К поверхности листьев и других органов растений грибница прикрепляется специальными присосками — аппрессориями. Обычно в цикле развития мучнисторосяных грибов имеются две стадии — конидиальная и сумчатая. Последняя возникает в результате полового процесса. Половые органы у мучнисторосяных грибов устроены довольно просто. Мужской (антеридий) состоит из двух клеток, женский (аскогон) — из одной. Содержимое верхней клетки антеридия переливается через специальное отверстие в женский орган и таким образом происходит оплодотворение. В результате его образуются аскоспоры (заключенные в сумки), находящиеся в замкнутых плодовых телах клейстотециях (клейстокарпиях). Освобождение аскоспор происходит после растрескивания клейстотеция. Споры выбрасываются наружу и, попадая на листья или другие органы восприимчивых растений, прорастают и вызывают их заражение. На поверхности зараженных органов развивается грибница, а на ней — конидиеносцы с конидиями (обычно по нескольку штук), расположенными в виде цепочек. У некоторых мучнисторосяных грибов в грибнице на конидиеносцах развивается лишь одна конидия. (рис. 86).

Конидии способны снова заражать растения, давая за лето несколько поколений. Таким образом, степень развития мучнисторосяных грибов зависит от того, на сколько благоприятны будут условия для развития конидиальной стадии. Как указывалось, грибница у большинства мучнисторосяных грибов поверхностная. Однако у некоторых родов этих грибов грибница или заходит в подустьичную полость (род Phyllactinia), или частично погружена в ткани растений (род Leveillula). В обоих случаях грибница развивается также на поверхности пораженных органов. Наличие погруженной в ткани грибницы — своеобразная защита гриба от высыхания. Грибы из рода левейллула (Leveillula) паразитируют на растениях в жарких засушливых местах.
На поверхности плодовых тел имеются особые выросты — придатки. По форме и расположению на плодовом теле придатков, а также по числу сумок в клейстотециях построена классификация мучнисторосяных грибов (рис. 87). Придатки могут быть простые, мало отличающиеся по форме от грибницы, разветвленные на концах или другой формы.

болезни: мучнистой росы поражает как озимую, так и яровую пшеницу, мучнистой росой картофель, Многие формы Erysiphe communis паразитируют на бобовых растениях и др.
28. Пиреномицеты
(Pyrenomycetes)
группа сумчатых грибов, плодовые тела которых — перитеции (См. Перитеций) — открываются на вершине порой или трещиной. Около 640 родов, объединяющих 6 тыс. видов (по другим данным, до 10 тыс. видов). Плодовые тела П. образуются или непосредственно на гифах мицелия, или на особом их сплетении — ложе, или строме, иногда внутри стромы. Спороношение в цикле развития сумчатое, а у многих П. и конидиальное, как у несовершенных грибов. Многие виды П. паразитируют в конидиальной стадии на высших растениях, затем после отмирания растений развиваются на них как сапрофиты и к весне образуют сумчатые спороношения, что обеспечивает грибу способность существования в самых разнообразных экологических условиях. Большинство П.— сапрофиты, живут преимущественно на растительных субстратах. Вместе с другими микроорганизмами П. играют роль в круговороте веществ. Большой вред наносят паразитические виды П. Например спорынья (Claviceps purpurea), склероции которой содержат алкалоиды, вызывает отравление человека и животных (эрготизм), Epichloё typhina — возбудитель чехловидной болезни злаков, некоторые виды Nectria, Eutypa, Nummularia, Diatrype, Valsa вызывают рак и некроз деревьев и кустарников, виды порядка Coryneliales — возбудители болезней многих тропических растений, Ceratocystis ulmi (или Ceratostomella ulmi) — возбудитель усыхания ильмовых, Calonectria graminicola — возбудитель снежной плесени озимых посевов, виды Phyllachora вызывают пятнистость листьев злаков. Виды Cordyceps паразитируют на личинках насекомых и грибах, на некоторых грибах живут и виды Nitschkia. К П. относятся широко используемые в биохимических и генетических исследованиях грибы Neurospora crassa и N. sitophila. Многие виды П.— микобионты лишайников.
29. Дискомицеты (лат. Discomycetes, нем. Diskomyzeten; греч. diskos — круг + mykēs (mykētos) — гриб)[1] — внетаксономическая группа (ранее — класс или порядок) сумчатых грибов[2], объединяющая несколько порядков (более 4 тысяч видов)… Плодовые тела у дискомицетов открытые (апотеции), большей частью блюдцевидные, с открыто расположенным сверху гимением. Большая часть видов дискомицетов относится к порядку Pezizales; их плодовые тела мясистые, часто ярко окрашенные. К порядку Hysteriales относятся Дискомицеты, имеющие чёрные вытянутые плодовые тела, раскрывающиеся узкой продольной щелью. Для дискомицетов порядка Phacidiales характерны закрытые плодовые тела, при созревании открывающиеся обычно лопастями вследствие разрыва сплетения гиф, покрывающего гимений. У дискомицетов из порядка Tuberales (трюфелевые) формируются подземные плодовые тела. Среди дискомицетов имеются сапрофиты и паразиты[3].Гимениальный слой, который состоит из сумок (аскусов) и парафиз, выстилает всю или только верхнюю поверхность плодового тела, бывшую внутренней, если плодовое тело в начале было замкнуто[2]. Размножение Предшествующий образованию сумок половой акт указывается для некоторых дискомицетов, однако по этому вопросу требуются ещё дальнейшие исследования[2]. Кроме плодоношения в сумках, существует ещё и конидиальное плодоношение (свободные конидиеносцы и конидиальные плоды); плодовые тела нередко развиваются на мицелии группами в большом числе, особенно у мелких форм[2]. У Д. из порядка Tuberales (трюфелевые) образуются подземные плодовые тела. Среди Д. имеются сапрофиты и паразиты. Особенно большой вред причиняют склеротинии, поражающие семечковые, косточковые, овощные и др. культурные растения. Съедобны трюфели, сморчки и строчки.
30. Loculoascomycetes - локулоаскомицеты. Для этих грибов характерны т.н. битуникатные, т.е. окруженные двойной оболочкой, аски. Наружная жесткая стенка (экзоаск, или экзотуника) при их созревании разрывается, через образовавшееся отверстие выпячивается внутренняя растяжимая стенка (эндоаск, или эндотуника), и только после этого в окружающую среду высвобождаются споры. Название класса связано с тем, что аски развиваются в полостях (локулах) внутри плодовых тел, обычно называемых аскостромами.Локулоаскомицеты выделены из числа плодосумчатых грибов в специальный подкласс на основании таких признаков, как двухслойная оболочка сумок и отсутствие у них настоящих плодовых тел. Сумки формируются в особых полостях — локулах, образующихся в мицелиальных стромах, которые получили название аскостром, или псевдотециев.Сумчатая стадия асколокулярных грибов развивается сапро-фитно на растительных остатках и обеспечивает первичное заражение растений. Паразитирующей и обеспечивающей дальнейшее распространение инфекции у асколокулярных грибов является конидиальная стадия.Среди многочисленных представителей подкласса следует выделить порядки плеоспрровые (Pleospcrales) и дотидейные (Dothideales). К первому из них относится гриб Venturia, являющийся сумчатой стадией возбудителей парши яблони и парши груши, ко второму — грибы рода Mycosphaerella и рода Didy-mella.К роду Mycosphaerella относятся сумчатые стадии таких: грибов, как Septoria, вызывающих белые пятнистости на груше. смородине, малине и других культурах, Phyllosticta — возбудитель бурой пятнистости листьев яблони, Ramularia — возбудитель белой пятнистости листьев земляники и др.К роду Didymella относят сумчатые стадии возбудителей аскохитоза огурца и аскохитоза гороха, пурпуровой пятнистости (дидимеллы) на малине и др.Псевдотеций Venturia, Didymella и Mycosphaerella, как правило, погружены в ткань питающего растения, имеют шаровидную или почти грушевидную форму, черные. Сумки обычно вытянутой формы — булавовидные, цилиндрические или почти грушевидные (немного «вздутые» в нижней части), часто согнутые, во многих случаях с псевдопарафизами (остатками межсумочной ткани), имеющими вид тонких нитей. Сумкоспоры двуклеточные, по созревании желтовато-зеленоватые или оливково-бурые.Строение и форма конидиальиых стадий у грибов, относящихся к локулоаскомицетам, весьма разнообразны, как и характер болезней, которые они вызывают. Так, конидии возбудителя парши яблони (Fusicladium dendriticum) имеют обратно-грушевидную форму и образуют на поверхности пораженных листьев пятна с зеленовато-бурым бархатистым налетом.У грибов родов Septoria и Phyllosticta конидиальное спороношение развивается в специальных споровместилищах с отвергшем на вершине — пикнидах (см. несовершенные грибы), форма и строение самих спор у них различны. У Septoria они вытянутые, почти нитевидные, с несколькими перегородками, а у Phyllosticta — овальные, одноклеточные.У гриба Ramularia tulasnei — возбудителя белой пятнистости листьев земляники — конидии цилиндрические, одноклеточные или с 1—2 перегородками, образуются на неветвящихся, собранных в пучки конидиеносцах.У гриба Ascochyta cucumis — возбудителя аскохитоза огурца и других тыквенных — конидии одно- или двуклеточные, развиваются в пикнидах.Конидиальные стадии асколокулярных грибов могут развиваться в течение вегетации в нескольких генерациях, вызывая новые, повторные заражения.У многих локулоаскомицетов конидиальные стадии способны перезимовывать на растительных остатках, не теряя своей жизнеспособности, и служить (наряду с сумчатой стадией) дополнительным источником первичной инфекции. На этом основании многих из них (Septoria, Ramularia, Ascochyta и др.) относят к классу несовершенных грибов.
31. базидиа́льные грибы́ —отдел царства грибов, включающий виды, производящие споры в булавовидных структурах, именуемых базидии. Вместе с аскомицетами относятся к подцарству высших грибов. Мицелий базидиомицетов септирован, каждая клетка содержит по два гаплоидных ядра. Обычно ядра расположены рядом посередине клетки, их пара носит название дикарион. Возле септы у гифов базидиомицетов формируется пряжка, участвующая при делении клетки. Жизненный цикл
Бесполое размножение базидиомицетов осуществляется конидиями, но происходит редко. Половое размножение происходит в виде соматогамии, при которой сливаются две вегетативные одноядерные клетки гаплоидного мицелия. У небольшого количества гомоталличных видов могут сливаться клетки одного и того же мицелия. Большинство видов являются гетероталличными, соответственно у них соматогамия происходит только между гифами с противоположными знаками «+» и «-». Половые органы у базидиальных грибов не образуются. зависимости от строения различают несколько типов базидий. Холобазидии имеют булавовидную форму и одноклеточное строение. Гетеробазидии состоят из расширенной нижней части — гипобазидии и верхней — эпибазидии, являющейся выростом гипобазидии. Фрагмобазидии, или телиобазидии, образуются из толстостенной покоящейся клетки путём образования поперечных перегородок, делящих её на четыре клетки. По бокам от этих клеток развиваются базидиоспоры. Представители
Согласно современным представлениям, отдел Basidiomycota делится на три подотдела и один класс, не включённый в подотдел. По предположению 2008 года, в мире насчитывается 16 классов, 52 порядков, 177 семейств, 1589 родов и 31515 видов Базидиомикота[2].Традиционно, отдел делился на два класса — Homobasidiomycetes, 1971 (включая шляпконожечные и др. грибы) и Heterobasidiomycetes R.T. Moore, 1980 (желеобразные, ржавчинные грибы, а также устомицеты). Ранее отдел Базидиомикота считался классом, и назывался Basidiomycetes Whittaker
32. Афиллофоровые грибы — второй по количеству видов порядок гименомицетов, не менее распространенный в природе, чем агариковые грибы. Внешний вид и микроскопическое строение плодовых тел афиллофоровых грибов очень варьируют как у представителей различных семейств, так нередко и у различных видов одного рода.Нетрудно отличить афиллофоровые грибы от всех других базидиомицетов: базидия у них всегда нераздельная, без перегородок; гименофор может иметь различные формы, но обычно не бывает пластинчатым, как у агариковых грибов, хотя есть исключения — некоторые трутовые грибы имеют пластинчатый гименофор, но в этом случае плодовые тела жесткокожистые или деревянистые (у агариковых плодовые тела, как правило, более или менее мясистые). Афиллофоровые грибы — сапрофиты, изредка паразиты травянистых растений и деревьев или микоризообразователи. Мицелий погружен в субстрат — в древесину, гумусовый слой почвы и т. п. Гифы мицелия хотя и многочисленные, но малозаметные, обыкновенно небольшого диаметра (2-10 мкм), бесцветные или слегка окрашенные, только у сравнительно малого количества видов образуют грибные шнурочки. У видов, обитающих в гниющей древесине, нередко образуются грибные пленки. Плодовые тела образуются обыкновенно через несколько месяцев или даже несколько лет после начала развития мицелия. В разнообразии форм плодовых тел можно различить пять типов: плодовые тела прямостоящие, шляповидные плодовые тела с центральной или эксцентрической ножкой., блюдцевидные или чашевидные плодовые тела, распростертые плодовые тела наименее заметные, шляповидные плодовые тела полукруглые, вееровидные, почковидные или продолговатые, прикрепленные боком или зачаточной боковой ножкой…. Около 4 тыс. видов; преимущественно сапротрофы. Возбудители гнилей древесины (некоторые трутовиковые), домовые грибы. Немногие афиллофоровые съедобны (лисички, некоторые ежовики и рогатики). Афиллофоровые грибы являяются разрушителями древесины и вызывают её гниение, могут наносить вред народному хозяйству.
33. Порядок Головневые(Ustilaginales) Головневые грибы — интересная во мпогих отношениях группа базидиомицетов. Известно около 1000 видов головневых грибов, объединенных в 40 родов. В СССР распространены более 300 видов. Головневые грибы поражают большинство органов своих растений-хозяев, в том числе вегетативные и цветочные почки, листья и стебли, лепестки венчика и чашелистики, тычинки и пестики, плоды и семена. Реже они поражают корни. Пораженные части растений выглядят как обугленные или покрытые сажей. Поэтому заболевания, вызываемые головневыми грибами, носят название «головня».Головневые грибы распространены от Арктики до тропиков, почти повсюду, где произрастают высшие растения, не исключая пустынь и гор. Споры головневых грибов чрезвычайно жизнеспособны и могут, прорастая, заразить высшее растение, даже пролежав в покое 20 и более лет. Например, отдельные споры возбудителя твердой головни пшеницы (Tilletia levis) сохраняют жизнеспособность на протяжении 25 лет, а споры возбудителя твердой головни ячменя (Ustilage hordei). головни кукурузы (Ustilago maydis, Головневые грибы гетероталличны: для образования нового поколения спор и для заражения высшего растения необходимо слияние двух внешне одинаковых, но физилогически различных баяидиоснор или мицелиальных клеток, относящихся к противоположным полам. Последние нередко обозначаются как + и —. У одних головневых грибов друг с другом сливаются базидиоспоры, находящиеся на базидий; у других — базидиоспоры, опавшие с базидий, или сливаются мицелии после прорастания базидиоспор; у третьих, у которых базидий не возникают, отдельные клетки фрагмобазидий обособлены перегородками. Половой процесс у головневых грибов состоит как бы из двух этапов. На первом из них происходит плазмогамия, т. е. слияние цитоплазмы двух раздельнополых клеток или гаплоидных мицелиев без слияния ядер. Плазмогамия происходит на покровной ткани растения-хозяина. На втором этапе в созревающих головневых спорах осуществляется кариогамия (слияние гаплоидных ядер). Головневые становятся диплоидными. Половой процесс у головневых грибов (впрочем, как и у других базидиомицетов) как бы растянут во времени и пространстве. От момента плазмогамии до момента кариогамии мицелий разрастается по клеткам, тканям и органам зараженного высшего растения. Естественно, каждая клетка такого мицелия двухъядерна. Заражение растений происходит двухъядерным мицелием, т. е. после полового процесса. Для успешной защиты одновременно следует проводить комплекс агротехнических приемов, улучшающих рост и развитие растений. Из них эффективны регулярные севообороты, соблюдение которых препятствует накоплению в почве спор возбудителей. Существенное влияние на степень поражения растений головневыми грибами оказывают сроки посева, глубина заделки семян, густота посева, способ уборки урожая, водный режим на полях и т. д.Существенным мероприятием по борьбе с головневыми заболеваниями растений является создание высокоустойчивых к болезням сортов сельскохозяйственных растений. Трудность выведения таких сортов состоит в том, что устойчивость к целому ряду заболеваний должна сочетаться с их высокой урожайностью. Лучшие советские сорта удовлетворяют этому требованию.
34.
35.
36. Пути заражения высших растений головневыми грибами различны. Основных путей четыре. Первый характеризуется тем, что головневые споры, удерживающиеся на семенах или сохраняющиеся в почве, прорастают на растении-хозяине, в частности на наклюнувшемся семени, еще до того, как образуются всходы. Этот путь заражения свойствен возбудителям твердой (мокрой) головни пшеницы (Tilletia caries, рис. 212), головни ржи (Tilletia secalis) и головни ячменя (Tilletia hordei), стеблевой головни ржи (Urocystis occulta) и стеблевой головни пшеницы (Urocystis tritici), пыльной головни кукурузы (Sorosporium reilianum,) и т. д. 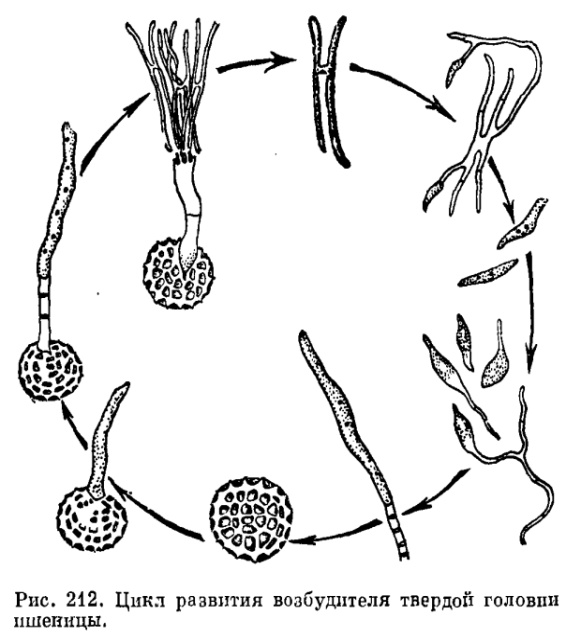
37. Для второго способа заражения, выявленного у возбудителей пыльной головни пшеницы (Ustilago tritici) и ячменя (Ustilago nuda), характерно прорастание головпевой споры на рыльце цветущего цветка, откуда гифы внедряются в развивающийся плод и сохраняются в виде мицелия в покоящемся состоянии в зародыше или оболочке семени до образования всходов в следующем году (рис. 213).

38. При третьем пути заражения, свойственном, в частности, возбудителю пузырчатой головни кукурузы (Ustilago maydis), головневые споры прорастают на молодых, начинающих рост и развитие надземных 33.-38. Порядок головнёвые - Ustilaginales. Головнёвые грибы - узкоспециализированные паразиты, вызывающие у растений болезни. известные под названием головня, для большинства из них характерен диффузный тип паразита - мицелий пронизывает все органы и ткани растения. Проявляются головневые чаще на репродуктивных органах - завязях, соцветиях. Органы растения, пораженные грибом, как правило, полностью разрушаются и превращаются в темную плотную порошащую массу, состоящую из огромного числа спор, носящих название устоспор (телиоспор) головневых спор. Этими спорами головневые распространяются и сохраняются. Прорастая, телиоспоры образуют базидию с базидиоспорами, которые у некоторых головневых грибов способны почковаться. Затем базидиоспоры попарно копулируют и, прорастая дикариотическим мицелием, заражают растения.
Головневые грибы паразитируют главным образом на зерновых - пшенице, ячмене, ржи, сорго, кукурузе, просе и др. Из овощных культур головней поражается, лук.
Название отдельных видов головни, как правило, связано с особенностями проявления внешнего вида. Пораженные части растения выглядят как обугленные или покрытые сажей. Поэтому заболевания, вызываемые головневыми грибами, носят название "головня": твердая, пыльная, стеблевая, пузырчатая и т.д.
В зависимости от строения базидий и расположения на ней базидиоспор. Базидиоспоры располагаются пучком на базидии. Порядок Головневые делят на два семейства – устилягиновые и тиллециевые.
В семейство устилягиновые – Ustilaginaсeae – объединены виды грибов с четырехклеточной базидией. На каждой клетке образуется по одной базидиоспоры. Наибольший интерес представляют 3 рода: устилаго, сороспориум и сфацелотека (рис. 16)
Для семейства тиллециевых – Tilletiaceae характерна одноклеточная базидия. Сюда относятся рода: тиллеция, уроцистис, энтилома.
Род энтилома – Entiloma. E.camissianum var. pratense заражает тимофеевку. E.dactylidis - ежу сборную.
По типам заражения (фазам, в которые происходит заражение, и источникам инфекции) все головневые грибы можно разделить на четыре основные группы.
1. Заражение растений происходит в период прорастания семян: а) за счет спор, находящихся на поверхности семян; б) за счет спор, находящихся в почве; в) за счет спор или кусочка мицелия, находящихся под пленкой (у пленчатых культур).
2. Заражение происходит в период цветения.
3. Заражение возможно в течение всей вегетации (в период активного роста растений).
4. Заражение происходит в период выходящего колоса. В таблице 7 приведен перечень основных видов головни на хлебных злаках и других культурах с указанием типа, по которому происходит заражение. Как видно из таблицы, у большинства видов головни заражение происходит в период прорастания семян за счет спор, находящихся на поверхности. Все эти виды проявляются уже в период созревания зерна, и телиоспоры попадают на семена во время обмолота (у твердой, карликовой, стеблевой, пузырчатой головни), либо несколько раньше, незадолго до уборки, если головневая масса может пылить в поле (пыльная головня овса, черная головня ячменя и др.).
заражение растений в период цветения происходит только у пыльной головни пшеницы и пыльной головни ячменя: проросток приникает в завязь. В период колошения колоски, пораженные этими видами головни, полностью разрушаются, превращаясь в пылящую головневую массу. Споры, рассеиваясь ветром, попадают на цветки здоровый растений (на рыльце пестика) и прорастают в фрагмобазидию (базидиоспоры у этих видов головни не образуются, а половой процесс осуществляется слиянием клеток самой базидии). Клетки фрагмобазидии попарно копулируют и, прорастая, формируют дикариотический мицелий, который и осуществляет заражение. Достигнув завязи, гриб приостанавливает свое развитие. Зерно формируется нормально и ни внешне, ни по товарным качествам не отличается от здорового, но для семенных целей оно уже непригодно. При посеве такого зерна разовьется больное растение, на котором весь колос, кроме центрального стержня, превратится в пылящую головневую массу в период цветения.
Из таблицы видно, что заражение растений возможно в течение всей вегетации, это характерно для пузырчатой головни кукурузы. Телиоспоры могут находиться на поверхности зерна или в почве. В течение всей вегетации заражаются молодые растущие ткани. На пораженных органах появляются вздутия вначале небольшие, позже увеличивающиеся в размерах. Сначала они покрыты беловатой пленкой, которая позже разрывается, обнаруживая пылящую черную массу спор.
В колосе поражаются не все зерна, а, как правило, бывает поражено 1-5 колосков. В незрелом колосе, не ломая его, трудно обнаружить индийскую головню, так как зараженные семена скрыты колосковыми чешуйками. При созревании больные колоски раскрываются больше здоровых, наружные и внутренние чешуйки раздвигаются и становятся видны зерна, пораженные головней. При небольшом сотрясении пораженные зерна выпадают из колосков, заражая почву.
Источником инфекции в поле являются телиоспоры, сохраняющиеся в почве или приставшие к поверхности здоровых семян.
Биологической особенностью возбудителя индийской головни является то, что телиоспоры прорастают после периода покоя (5-6 мес) и заболевание в первый год может не проявиться, но споры могут сохраняться жизнеспособными в почве больше 4 лет. Иногда болезнь заметна в год высева. Для РФ индийская головня - карантинный объект.
вегетативных и репродуктивных органах своего хозяина, куда они проникают.
39. Порядок Ржавчинные (Uredinales).Грибы, относящиеся к этому порядку, вызывают заболевания многих растений. Симптомы поражения растений — пятна или полосы обычно ржаво-бурого цвета. Ржавчинные грибы паразитируют на высших сосудистых растениях из многих семейств. Отличительной чертой этой группы грибов является наличие в их цикле развития нескольких различных по форме и функциям спороношений. Эти спороношения называются стадия ми развития гриба. Разные виды спор могут развиваться как на одном растении (однохоэяинные виды), так и на разных растениях (разнохозяинные виды).При развитии ржавчины на растении обычно происходит местное, локальное заражение, т. е. поражение паблюдается па небольшом участке, куда попала одна спора гриба. У небольшого числа видов мицелий может пронизывать все растение и давать так называемое диффузное поражение всего растения. Для ржавчинных грибов характерно также массовое рассеивание спор. Оседая на растениях, споры ржавчины, переносимые ветром на многие километры, вызывают множество местных очагов инфекции на растении одного и того же вида. Порядок ржавчинных грибов делят на два семейства — мелампсоровых (Melampsoraceae) и пукциниевых (Pucciniaceae). Прорастающая спора ржавчинного гриба дает так называемую ростковую трубку, которая чаще всего через устьица проникает в ткани растения-хозяина. Мицелий гриба растет и распространяется внутри тканей растения, посылая в клетки гаустории, с помощью которых происходит питание гриба. Гифы ржавчинных грибов распространяются по межклетникам растения-хозяина. Контакт между цитоплазмой хозяина и паразита осуществляется в кончиках гаусторий. Недавно было показано, что на конце гаустории, примыкающей к цитоплазме растения-хозяина, оболочки нет. В этом месте есть только цитоплазменная мембрана, через которую питательные вещества хозяина переходят в мицелий паразита. Важной особенностью ржавчинных грибов является приуроченность их к паразитированию на определенных растениях-хозяевах. Например, Puccinia graminis состоит из отдельных специализированных форм, которые различаются в основном по паразитированию на отдельных родах семейства злаков. Известны специализированные формы, приуроченные к пшенице (стеблевая ржавчина пшеницы), ржи (стеблевая ржавчина ржи), овсу (стеблевая ржавчина овса). Некоторые специализированные формы поражают дикорастущие злаки — полевицу, мятлик, тимофеевку, вейник и др.