Синапс (греч. synapsis — соединение) — это специализированная структура, обеспечивающая передачу сигнала от клетки к клетке. Посредством синапса реализуется действие многих фармакологических препаратов.
Структурно-функциональная организация. Каждый синапс имеет пре - и постсинаптическую мембраны и синаптическую щель (рис. 17).
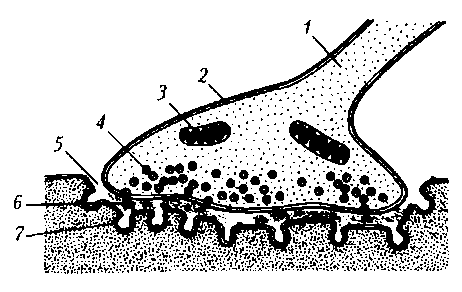
Рис. 17. Нервно-мышечный синапс скелетной мышцы: 1 – ветвь аксона; 2 – пресинаптическое окончание аксона; 3 – митохондрия; 4 – синаптические пузырьки, содержащие ацетилхолин; 5 – синаптическая щель; 6 – молекулы медиатора в синаптической щели; 7 – постсинаптическая мембрана мышечного волокна с N-холинорецепторами
Пресинаптическая мембрана нервно-мышечного синапса представляет собой часть мембраны пресинаптического окончания аксона мотонейрона. Через нее осуществляется выброс (экзоцитоз) медиатора (лат. mediator — посредник) в синаптическую щель. В нервно-мышечном синапсе медиатором является ацетилхолин. Медиатор пресинаптического окончания содержится в синаптических пузырьках (везикулах), диаметр которых составляет около 40 нм. Они образуются в комплексе Гольджи, с помощью быстрого аксонного транспорта доставляются в пресинаптическое окончание, где заполняются медиатором и АТФ. В пресинаптическом окончании содержится несколько тысяч везикул, в каждой из которых имеется от 1 тыс. до 10 тыс. молекул химического вещества.
Постсинаптическая мембрана (концевая пластинка в нервно-мышечном синапсе) — это часть клеточной мембраны иннервируемой мышечной клетки, содержащая рецепторы, способные связывать молекулы ацетилхолина. Особенность этой мембраны: множества мелких складок, увеличивающих ее площадь и количество рецепторов на ней до 10—20 млн в одном синапсе.
Синаптическая щель в нервно-мышечном синапсе имеет ширину в среднем 50 нм. Она содержит межклеточную жидкость, ацетилхолинэстеразу и мукополисахаридное плотное вещество в виде полосок, мостиков, в совокупности образующих базальную мембрану, соединяющую пре- и постсинаптическую мембраны.
Механизмы синаптической передачи включают три основных этапа (рис. 18).

Рис. 18. Механизм проведения импульса через химический синапс: 1-8 – этапы процесса (Чеснокова, 2007)
Первый этап — процесс выброса медиатора в синаптическую щель, который запускается посредством ПД пресинаптического окончания. Деполяризация его мембраны ведет к открытию потенциалуправляемых Са-каналов. Са2+ входит в нервное окончание согласно электрохимическому градиенту. Часть медиатора в пресинаптическом окончании локализуется на пресинаптической мембране изнутри. Са2+ активирует экзоцитозный аппарат пресинапса, представляющий собой совокупность белков (синапсин, спектрин и др.), пресинаптического окончания, активация которых обеспечивает выброс ацетилхолина посредством экзоцитоза в синаптическую щель. Количество высвобождаемого ацетилхолина из пресинаптического окончания пропорционально в четвертой степени количеству поступившего туда Са2+. На один ПД из пресинаптического окончания нервно-мышечного синапса выбрасывается 200—300 квантов (везикул) медиатора.
Второй этап — диффузия ацетилхолина в течение 0,1—0,2 мс к постсинаптической мембране и действие его на N-холинорецепторы (стимулируются также никотином, вследствие чего и получили свое название). Удаление ацетилхолина из синаптической щели осуществляется путем разрушения его под действием ацетилхолинэстеразы, расположенной в базальной мембране синаптической щели, в течение нескольких десятых долей миллисекунды. Около 60% холина захватывается обратно пресинаптическим окончанием, что делает синтез медиатора более экономичным, часть ацетилхолина рассеивается. В промежутках между ПД из пресинаптического окончания происходит спонтанное выделение 1— 2 квантов медиатора в синаптическую щель в течение 1 с, формируя так называемые миниатюрные потенциалы (0,4—0,8 мВ). Они поддерживают высокую возбудимость иннервируемой клетки в условиях функционального покоя и выполняют трофическую роль, а в ЦНС — способствуют поддержанию тонуса ее центров.
Третий этап — взаимодействие ацетилхолина с N-холинорецепторами постсинаптической мембраны, в результате чего открываются ионные каналы на 1 мс и, вследствие преобладания входа N+ в клетку, происходит деполяризация постсинаптичедкой мембраны (концевой пластинки). Эту деполяризацию в нервно-мышечном синапсе называют потенциалом концевой пластинки (ПКП) (рис. 19).
Особенностью нервно-мышечного синапса скелетного мышечного волокна является то, что при одиночной его активации формируется ПКП большой амплитуды (30—40 мВ), электрическое поле которого вызывает генерацию ПД на мембране мышечного волокна вблизи синапса. Большая амплитуда ПКП обусловлена тем, что нервные окончания делятся на многочисленные веточки, каждая из которых выбрасывает медиатор.
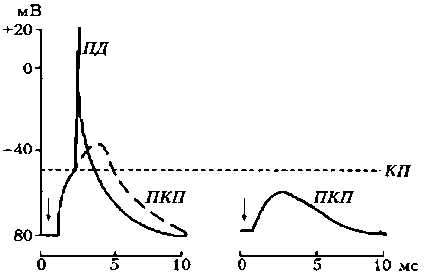
Рис. 19. Потенциал концевой пластинки (Шмидт, 1985): КП – критический потенциал; ПД – потенциал действия; А – ПКП в нормальной мышце; Б – ослабленный ПКП в курарезированной мышце; стрелками указан момент нанесения стимула
Характеристика проведения возбуждения в химических синапсах. Одностороннее проведение возбуждения от нервного волокна к нервной или эффекторной клетке, так как пресинаптическое окончание чувствительно только к нервному импульсу, а постсинаптическая мембрана — к медиатору.
Неизолированное — возбуждение рядом расположенных постсинаптических мембран суммируется.
Синаптическая задержка в передаче сигнала к другой клетке (в нервно-мышечном синапсе 0,5-1,0 мс), что связано с высвобождением медиатора из нервного окончания диффузией его к постсинаптической мембране и возникновением постсинаптических потенциалов, способных вызвать ПД.
Декрементность (затухание) возбуждения в химических синапсах при недостаточном выделении медиатора из пресинаптических окончаний в синаптические щели.
Низкая лабильность (в нервно-мышечном синапсе составляет 100 Гц), которая в 4 - 8 раз ниже лабильности нервного волокна. Это объясняется синаптической задержкой.
Проводимость нервно-мышечного синапса (как и химических синапсов ЦНС) угнетается или, наоборот, стимулируется различными веществами.
Например, кураре и курареподобные вещества (диплацин, тубокурарин) обратимо связываются с N-холинорецепторами постсинаптической мембраны, блокируют действие на нее ацетилхолина и передачу в синапсе. Напротив, некоторые фармакологические препараты, например прозерин, подавляют активность ацетилхолинэстеразы, способствуя умеренному накоплению ацетилхолина и облегчению синаптической передачи, что используется в лечебной практике.
Утомляемость (синаптическая депрессия) — ухудшение проводимости вплоть до полной блокады проведения возбуждения при длительном функционировании синапса (главная причина — истощение медиатора в пресинаптическом окончании).
Вопросы для самоконтроля
1.Каков механизм распространения возбуждения по нервному волокну? Какова роль перехватов Ранвье в проведении возбуждения по миелинизированному нервному волокну?
2.В чем преимущество скачкообразного (сальтаторного) распространения возбуждения над непрерывным его проведением вдоль мембраны волокна?
3.В чем физиологическое значение изолированного проведения возбуждения по нервному волокну?
4.Какие нервные волокна (афферентные или эфферентные, вегетативные или соматические) относятся к группе А? Какова скорость проведения возбуждения по ним?
5.Какие нервные волокна (афферентные или эфферентные, вегетативные или соматические) относятся к группе В? Какова скорость проведения по ним?
6.Какие нервные волокна (афферентные или эфферентные, вегетативные или соматические) относятся к группе С? Какова скорость проведения возбуждения по ним?
7.Перечислите структуры нервно-мышечного синапса (скелетная мышца). Что называют концевой пластинкой?
8.Перечислите последовательность процессов, ведущих к освобождению медиатора из пресинаптической мембраны в синаптическую щель при передаче возбуждения в синапсе.
9. Локальным потенциалом или распространяющимся возбуждением является потенциал концевой пластинки?
10.Что такое миниатюрные потенциалы концевой пластинки, каков механизм их возникновения?
11.В чем заключается трофическое влияние нерва на мышцу, осуществляемое через нервно-мышечный синапс?
12.Какие вещества являются медиаторами в нервно-мышечных синапсах гладкой и поперечнополосатой мышц?
13.Что такое сенсорный рецептор?
14.На какие две группы делятся сенсорные рецепторы по скорости адаптации? Назовите рецепторы, относящиеся к каждой из них.
15.Что понимают под первичными и вторичными рецепторами?
16.Перечислите основные свойства рецепторов.
17.Что называют адаптацией рецепторов? Как изменяется частота импульсов в афферентном нервном волокне при адаптации рецептора?
18.Назовите локальные потенциалы, возникающие при возбуждении первичных и вторичных рецепторов.
19.Рецепторный потенциал, где он возникает, каково его значение?
20.Генераторный потенциал, где он возникает, каково его значение?
21.Где возникает потенциал действия при возбуждении первичного сенсорного рецептора?
22. Где возникает потенциал действия при возбуждении вторичного сенсорного рецептора?
Физиология мышц
1.3.1. Структурнофункциональная характеристика скелетной мышцы
Мышцы подразделяют на поперечнополосатые (скелетная и сердечная) и гладкие (сосуды и внутренние органы, кроме сердца).
Скелетная мышца состоит из мышечных волокон, изолированных в структурном и функциональном отношении друг от Друга, которые представляют собой вытянутые многоядерные клетки. Толщина волокна составляет 10—100 мкм, а его длина варьирует в пределах от нескольких миллиметров до нескольких сантиметров. Количество мышечных волокон, установившись постоянным на 4—5-м месяце постнатального онтогенеза, в последующем не изменяется; с возрастом изменяются (увеличиваются) лишь их длина и диаметр.
Назначение основных структурных элементов. Характеристика основных элементов мышечного волокна. От клеточной мембраны мышечного волокна (сарколеммы) вглубь отходят многочисленные поперечные инвагинации (Т-трубочки), которые обеспечивают ее взаимодействие с саркоплазматическим ретикулулом (СПР) (рис. 20).

Рис. 20. Взаимоотношение клеточной мембраны (1), поперечных трубочек (2), боковых цистерн (3) и продольных трубочек (4) саркоплпзматическаого ретикулума, сократительных белков (5): А – в состоянии покоя; Б – при сокращении мышечного волокна; точками обозначены ионы Ca2+
СПР представляет собой систему связанных друг с другом цистерн и отходящих от них в продольном направлении канальцев, расположенных между миофибриллами. Терминальные (концевые) цистерны СПР примыкают к Т-трубочкам, формируя так называемые триады. В цистернах содержится Са2+, играющий важную роль в мышечном сокращении. В саркоплазме имеются внутриклеточные элементы: ядра, митохондрии, белки (в том числе миоглобин), капельки жира, гранулы гликогена, фосфатсодержащие вещества, различные малые молекулы и электролиты.
Миоибриллы — субъединицы мышечного волокна. В одном мышечном волокне может насчитываться более 2 тыс. миофибрилл, их диаметр 1—2 мкм. В одиночной миофибрилле содержится 2—2,5 тыс. протофибрилл — параллельно расположенных нитей белка (тонкие — актин, толстые — миозин). Актиновые нити состоят из двух субъединиц, скрученных в виде спирали. В состав тонких нитей входят также регуляторные белки — тропомиозин и тропонин (рис. 21).
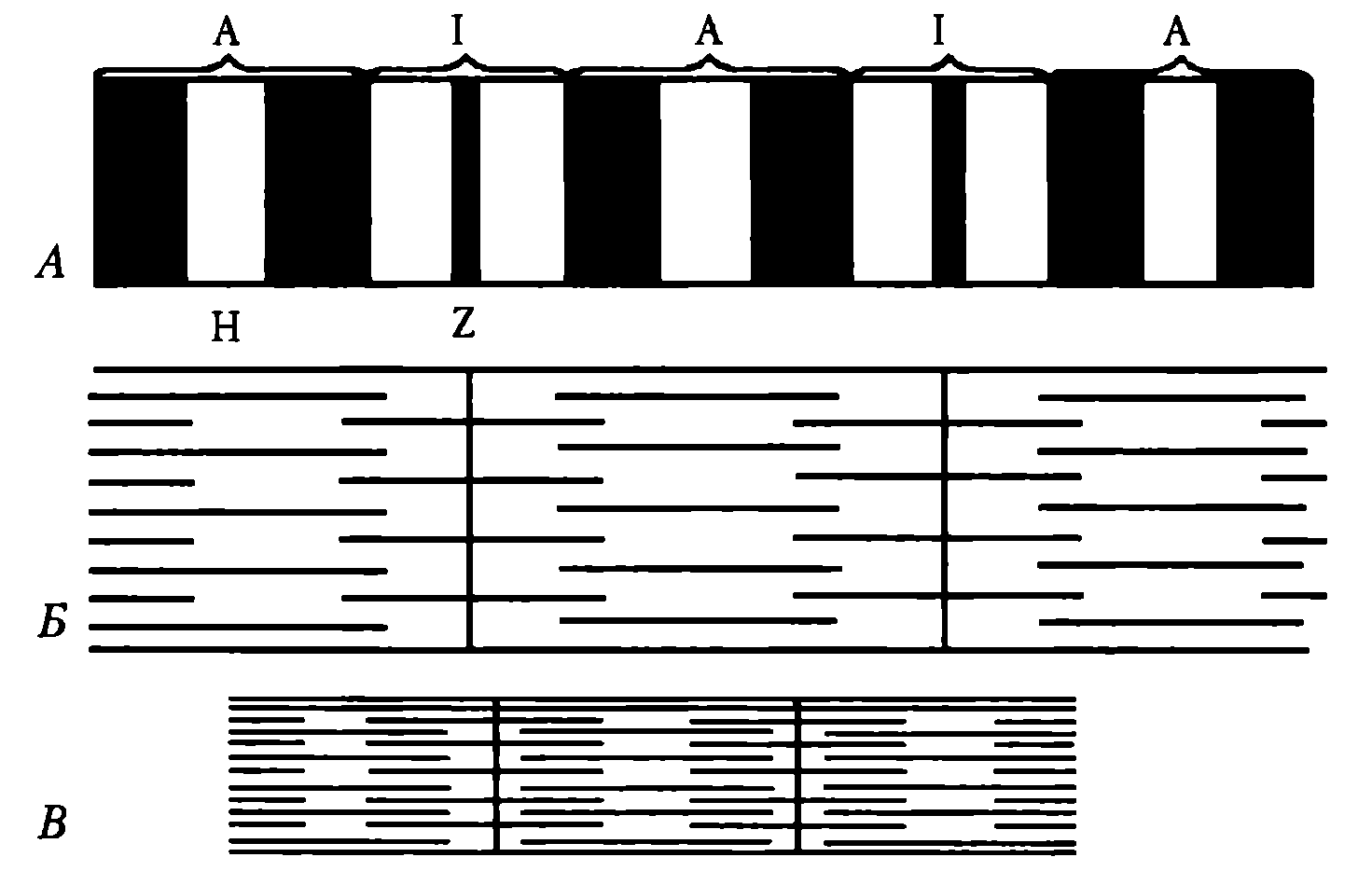
Рис. 21. Взаимное расположение структурных элементов миофибрилл при их расслаблении (А,Б) и сокращении (В)
Эти белки в невозбужденной мышце препятствуют взаимосвязи актина и миозина, поэтому мышца в покое находится в расслабленном состоянии. Миофибриллы включают в себя последовательно соединенные блоки — саркомеры (Б), отделенные друг от друга Z-полосками. Саркомер (длина 2—Змкм) является сократительной единицей мышечного волокна; при длине 5см оно включает в себя около 20 тыс. последовательно соединенных саркомеров. Миофибриллы отдельного мышечного волокна связаны таким образом, что расположение саркомеров совпадает, и это создает картину поперечной исчерченности волокна при наблюдении в световом микроскопе (рис. 22).

Рис. 22. Саркомер миоцита скелетной мышцы (A. Vander, J. Sherman, D. Luciano, 2004)
Элементы саркoмера (см. рис. 21). Миозиновые протофибриллы образуют наиболее темную часть саркомера — А-диск (анизотропный, он сильно поляризует белый свет). Более светлый участок в центре А-диска называют Н-зоной. Светлый участок саркомера между двумя А-дисками называют 1-диском (изотропный, почти не поляризует свет). Он образован актиновыми протофибриллами, идущими в обе стороны от Z-полосок. Каждый саркомер имеет два набора тонких нитей, прикрепленных к Z-полоскам, и один комплект толстых нитей, сосредоточенных в А-диске. В расслабленной мышце концы толстых и тонких филаментов в разной степени перекрывают друг друга на границе между А- и 1-дисками.
Классификация мышечных волокон:
По структурно-функциональным свойствам и цвету выделяют две основные группы мышечных волокон: быстрые и медленные.
Белые (быстрые) мышечные волокна содержат больше миофибрилл и меньше — митохондрий, миоглобина и жиров, но больше гликогена и гликолитических ферментов; эти волокна называют гликолитическими. Капиллярная сеть, окружающая эти волокна, относительно редкая. Скорость рабочего цикла у данных волокон примерно в 4 раза больше, чем у медленных, что объясняется более высокой АТФазной активностью быстрых волокон, но они обладают малой выносливостью. У белых мышечных волокон число нитей актина и миозина больше, чем у красных, поэтому они толще и сила их сокращения больше, чем у красных волокон.
Красные мышечные волокна содержат много митохондрий, миоглобина, жирных кислот. Эти волокна окружены густой сетью кровеносных капилляров, они имеют меньший диаметр. Митохондрии обеспечивают высокий уровень окислительного фосфорилирования, поэтому данные волокна называют оксидативными. Красные мышечные волокна подразделяются на две подгруппы: быстрые и медленные. Медленные волокна могут выполнять работу в течение относительно продолжительного периода времени; утомление в них развивается медленнее. Они более приспособлены к тоническим сокращениям. Красные быстрые волокна по скорости утомления занимают промежуточное положение между белыми и красными медленными. Скорость их сокращения близка к скорости сокращения белых волокон, что также объясняется высокой АТФазной активностью миозина красных быстрых волокон.
Также имеется незначительное число истинных тонических мышечных волокон; на них локализуется по 7-10 синапсов, принадлежащих, как правило, нескольким мотонейронам, например, в глазодвигательных мышцах, мышцах среднего уха. ПКП этих мышечных волокон не вызывают генерации ПД в них, а непосредственно запускают мышечное сокращение.
Группа мышечных волокон, двигательную (нейромоторную) единицу. В мышцах, совершающих быстрые и точные движения, например в глазодвигательных, нейромоторные единицы состоят из 3—5 мышечных волокон. В мышцах, осуществляющих менее точные движения (например, мышцы туловища и конечностей), двигательные единицы включают сотни и тысячи мышечных волокон. Большая двигательная единица, по сравнению с малой, включает крупный мотонейрон с относительно толстым аксоном, который образует большое число концевых веточек в мышце и, следовательно, иннервирует большое число мышечных волокон. Все мышечные волокна одной двигательной единицы, независимо от их количества, относятся к одному типу. Все скелетные мышцы по своему составу являются смешанными, т.е. образованы красными и белыми мышечными волокнами.
Специфическим свойством всех мышц является сократимость — способность сокращаться, т.е. укорачиваться или развивать напряжение. Реализация этой способности осуществляется с помощью возбуждения и его проведения по мышечному волокну (свойства соответственно возбудимости и проводимости).
Скелетные мышцы не обладают автоматией, управляются организмом произвольно импульсацией из ЦНС, поэтому их называют также произвольными. Гладкие мышцы по собственному желанию не сокращаются, поэтому их называют также непроизвольными, но они обладают автоматией.
Функции скелетной мышцы:
Обеспечение двигательной активности организма — поиск и добывание воды и пищи, ее захват, жевание, глотание, оборонительные реакции, трудовая деятельность — физическая и творческая работа художника, писателя, ученого, композитора в конечном итоге выражается в движении: рисование, письмо, игра на музыкальном инструменте и т.п.
Обеспечение дыхания (движений грудной клетки и диафрагмы).
Коммуникативная функция (устная и письменная речь, мимика и жесты).
Участие в процессах терморегуляции организма с помощью изменения интенсивности сократительного термогенеза.