Регуляция экспрессии генов с помощью антисмысловых РНК характеризуется высокой специфичностью. Это обусловлено большой точностью процесса РНК-РНК-гибридизации, основанной на комплементарном взаимодействии друг с другом протяженных последовательностей нуклеотидов. Однако сами по себе антисмысловые РНК не инактивируют необратимо мРНК-мишени, и для подавления экспрессии соответствующих генов требуются высокие (по крайней мере, эквимолярные по отношению к мРНК) внутриклеточные концентрации антисмысловых РНК. Эффективность действия антисмысловых РНК резко повысилась после того, как в их состав были введены молекулы рибозимов – коротких последовательностей РНК, обладающих эндонуклеазной активностью. Хотя ниже термин "рибозимы" будет использоваться почти исключительно в отношении молекул РНК, обладающих эндонуклеазной активностью, необходимо иметь в виду, что в настоящее время известно множество других ферментативных активностей, ассоциированных с РНК. Поэтому рибозимами в широком смысле называют молекулы РНК, обладающие любой ферментативной активностью. Некоторые из этих активностей будут рассмотрены в разделе 9.3 при обсуждении возможной роли РНК в происхождении жизни.
Типы рибозимов
Эндорибонуклеазная активность РНК была впервые обнаружена Т. Чехом в 1980 г. у интрона группы I предшественника рибосомной РНК Tetrahymena, осуществляющего аутокаталитическую реакцию сплайсинга (аутосплайсинга), в результате которой происходит вырезание из молекулы предшественника рРНК последовательности этого интрона и образование зрелой молекулы рРНК. (Молекулярный механизм реакции аутосплайсинга уже был рассмотрен в первой части книги в разделе 3.3.2.) С тех пор аутокаталитические реакции расщепления были выявлены у многих молекул РНК (рис. II.25). Наиболее перспективными для использования в составе антисмысловых РНК оказались рибозимы, образующие пространственную структуру типа "головки молотка" (hammerhead – HH) (см. рис. II.25, а). Такие РНК были найдены у сателлитных РНК вирусов растений, вироидов, а также среди транскриптов сателлитных ДНК тритонов. У РНК подобного типа была определена минимальная каноническая последовательность, необходимая для появления ферментативной активности, длиной в 27 нуклеотидов, из которых высококонсервативны 13 нуклеотидов, а также последовательность из трех нуклеотидов, в пределах которой происходит расщепление фосфодиэфирной связи (см. рис. II.25, а).
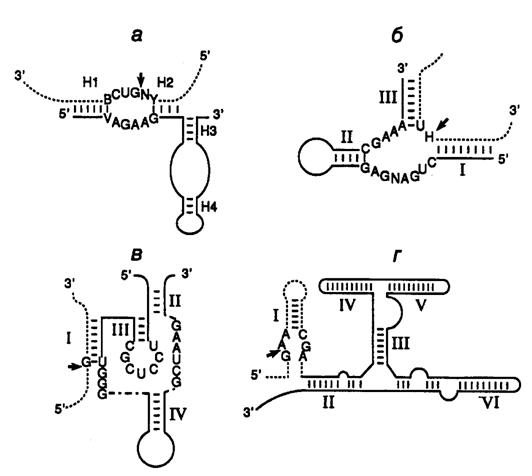
Рис. II.25. Особенности вторичной структуры рибозимов разных типов
а – "головка молотка", б – шпилька, в – рибозим вируса гепатита d, г – рибозим Neurospora VS. Стрелки обозначают точки расщепления РНК; нуклеозид N может быть A, U, G или C, H – A, U или C, Y – любой пиримидин; приведены общепринятые обозначения элементов вторичных структур рибозимов
Другими примерами рибозимов являются РНК, образующие структуры типа шпилек, а также РНК вируса гепатита δ (ВГД) и сателлитная РНК Варкуда (Varcud satellite – VS) нейроспоры (см. рис. II.25, б,в,г соответственно). (Структура и механизм действия рибозимов в составе интронов групп I, II и III были подобно рассмотрены в разделе 2.2.3.) Рибозимы со структурами типа шпилек были впервые обнаружены у вироидов и вирусоидов растений, рибозимы ВГД были идентифицированы среди сателлитных РНК вируса гепатита В человека, а рибозимы VS – среди сателлитных РНК природных изолятов Neurospora. В связи с рибозимами следует вспомнить и о РНКазе Р E. coli, состоящей из небольшого полипептида и РНК длиной в 375 нуклеотидов, о которой уже упоминалось в разделе 2.2.1. Именно РНК обеспечивает этому ферменту эндонуклеазную активность, необходимую для процессинга предшественников тРНК.
Хотя у природных РНК сайт расщепления и последовательность нуклеотидов рибозима расположены на одной молекуле (in cis по отношению друг к другу), HH-рибозимы могут действовать и на сайты расщепления, расположенные in trans, т.е. на других молекулах РНК. Именно это свойство рибозимов данного класса позволило использовать их для создания высокоспецифичных эндорибонуклеаз на основе антисмысловых РНК, которые обладают способностью расщеплять in trans молекулы РНК, содержащие тринуклеотидные сайты расщепления. Для создания высокоспецифического рибозима, функционирующего по этому принципу, последовательность рибозима заключают в последовательность антисмысловой РНК таким образом, чтобы при образовании гибрида с РНК-мишенью в контакте с активным центром рибозима оказывалась тринуклеотидная последовательность сайта расщепления.
Свойства рибозимов
Стабильность рибозимов в биологических жидкостях. Нестабильность РНК является одним из основных ограничений, препятствующих эффективному их использованию in vivo в качестве лекарственных средств. Поскольку рибозимы представляют собой короткие молекулы, которые можно получать в результате химического синтеза, химические модификации этих олигонуклеотидов рассматриваются как одна из перспективных возможностей повышения их устойчивости к нуклеазам. Было установлено, что введение 2’- O- Me-модифицированных нуклеотидов вместо обычных во все, кроме пяти, положения HH-рибозима позволило повысить его стабильность, по крайней мере, в 1000 раз без существенной потери каталитической активности. Другие модификации нуклеотидов в определенных положениях: введение 2'-О-аллильных групп, фосфоротиоатных связей вместо фосфодиэфирных, замена пиримидинов их 2’-амино- или 2’-фторпроизводными, использование различных 2’-производных сахаров в сахарофосфатном остове РНК и т.п., как правило, приводили к снижению активности рибозимов, но сопровождались повышением их стабильности. Улучшенные конструкции рибозимов с модифицированными сахарами обладали каталитической активностью, свойственной рибозимам дикого типа, со временем полужизни в сыворотке крови человека 5–8 ч. Кроме того, введение на 3’-конец рибозима остатка dT, соединенного связью 3’-3’, сопровождалось увеличением времени его полужизни в сыворотке до 3 дней.
Адресная доставка искусственных рибозимов. Существуют два основных подхода к адресной доставке олигонуклеотидов. При одном из них синтезированные молекулы рибозимов объединяют химическими методами или физически с макромолекулами, например липидами или лигандами рецепторов. В зависимости от природы молекул, находящихся в комплексе с рибозимом, его доставка будет либо неспецифической (в составе липосом), либо более специфической, которая может обеспечиваться, например эндоцитозом, опосредованным рецепторами. Другая группа методов использует экспрессирующие векторы, содержащие ген рибозима, транскрипция которого сопровождается внутриклеточным биосинтезом его молекул.
В ходе исследований по оптимизации адресной доставки рибозимов были изучены многие конструкции липидных носителей. Такие конструкции позволяют инкапсулировать большое количество молекул рибозимов как в водном окружении, так и растворенными в липидном бислое. Соединение липосом с молекулами соответствующих антител или других лигандов может до некоторой (хотя и небольшой) степени обеспечивать тканеспецифическую доставку рибозимов и их проникновение в клетки требуемых типов, например пораженных вирусами. Циркулирующие липосомы быстро удаляются из кровотока макрофагами ретикулоэндотелиальной системы. Химическая модификация липидного бислоя или покрытие липосом оболочкой из полиэтиленгликоля ослабляют неспецифическую сорбцию белков сыворотки, а следовательно, и неспецифическое распознавание липосом макрофагами.
Для создания альтернативных гидрофобных комплексов молекул рибозимов с носителем иногда используют катионные липиды. Такие липиды содержат длинные цепи жирных кислот (обычно C16 или C18), соединенные с полиамином. Полагают, что комплексы катионных липидов с олигонуклеотидами не образуют липосомоподобных структур. Результаты, полученные при исследовании возможностей адресной доставки рибозимов с помощью липосом и другими аналогичными методами, вселяют оптимизм. Однако до сих пор эти методы остаются чисто эмпирическими и должны разрабатываться индивидуально для каждого нового рибозима и типа клеток, в которые предполагается его доставлять.
Для адресной доставки генов рибозимов в составе экспрессирующих векторов используют те же приемы, что и при генотерапии (подробнее см. раздел 10.4). Особенно часто для этой цели применяются ретровирусы. Включение в состав рибозима сигнальной последовательности этих вирусов, обеспечивающей упаковку их РНК в вирусные частицы, на 90% снижает титр соответствующих ретровирусов. При этом рибозим не оказывал влияния на те же мишени, локализованные в цитоплазме.
Аденоассоциированные вирусы (ААВ), способные доставлять свою ДНК в клетки многих типов, также часто используются в качестве носителей для векторов, экспрессирующих рибозимы. Недавно сконструирован вектор, который из всех последовательностей ААВ содержал только концевые последовательности. С использованием этого вектора получена экспрессия антисмысловых РНК гена tar вируса иммунодефицита человека, которая блокировала его размножение. С помощью аденовирусов можно не только направленно осуществлять доставку экспрессирующихся генов, но и переносить большие молекулы декстранов, белков и плазмид, связанных с лигандами, способными и неспособными к независимой репликации. Например, удается транспортировать гены к дыхательному эпителию в составе комплексов аденовирус–полилизин–ДНК. Адресная доставка векторов к требуемым клеткам или тканям может быть осуществлена также непосредственно в виде аэрозоля (эпителий легких) или ex vivo в случае клеток костного мозга, который далее повторно вводят в организм больного.
Исследование функционирования рибозимов in vivo. Для доказательства возможности использования рибозимов как лекарственных средств необходимо было прежде всего продемонстрировать, что осуществляемое ими расщепление мРНК сопровождается ожидаемыми физиологическими и биохимическими изменениями в организме. В одних из первых опытов использовали рибозимы, направленные против мРНК хлорамфениколацетилтрансферазы (ХАТ), ген которой экспрессировался в клетках животных. Рибозимы, синтез которых обеспечивался экспрессирующими векторами, специфически на 60% подавляли внутриклеточный синтез ХАТ. Кроме того, было показано направленное действие рибозимов против 28S рРНК и ее предшественников, а также мРНК a-лактальбумина, но не других родственных РНК, находящихся в клетках. В результате в настоящее время считается признанным, что принцип действия рибозимов подтвержден на клеточном уровне.
Для доказательства эффективности действия рибозимов in vivo с помощью этих агентов была предпринята попытка индуцировать диабет у мышей путем воздействия на мРНК глюкокиназы b-клеток поджелудочной железы с помощью специфически экспрессирующегося в них трансгена рибозима. Оказалось, что у экспериментальных мышей активность глюкокиназы снижена в три раза именно в b-клетках, что приводило к нарушению секреции ими инсулина и ответа на глюкозу. При этом рибозим не оказывал влияния на функционирование клеток печени. Тем не менее уровень глюкозы в крови таких трансгенных мышей оставался прежним, что позволило сделать вывод о важности дефицита по глюкокиназе печени, а не только поджелудочной железы для развития симптомов заболевания. Эти опыты продемонстрировали возможность использования рибозимов для исследования физиологии у интактных животных.
Эффектные результаты с рибозимами удалось получить и на дрозофиле. Были созданы трансгенные яйца мух, содержащие ген рибозима, действующего на мРНК гена fushi tarazu (ftz), находящегося под контролем температурно-чувствительного промотора. Этот гомеозисный ген уже упоминался в связи с обсуждением регулирующих его экспрессию факторов транскрипции в первой части книги. Индуцируя рибозим в определенные фазы эмбрионального развития дрозофилы, установили, что вначале нарушение функционирования гена ftz приводит к нарушению сегментации кутикулы у личинок. Если ген инактивировали во время нейрогенеза, то нарушалось развитие центральной нервной системы на фоне нормального развития кутикулы. Эти опыты показали, что индукция рибозима у дрозофилы фенотипически проявляется точно так же, как мутации, инактивирующие ген ftz,и не сопровождается другими видимыми нарушениями в организме.
Введение в мужские пронуклеусы оплодотворенных ооцитов мышей плазмиды, экспрессирующей рибозим, направленный против мРНК b2-макроглобулина, сопровождалось его последующим синтезом в легких, почках и селезенке взрослых животных. При этом в легких происходило снижение уровня мРНК b2-макроглобулина более чем на 90%. Не столь значительное уменьшение содержания мРНК наблюдали в почках и селезенке.
Первые опыты с рибозимами, включенными в состав антисмысловых РНК, показали их высокую эффективность и специфичность in vitro и меньшую эффективность in vivo. Это может быть связано с тем, что РНК в эукариотических клетках, как правило, находится в составе больших рибонуклеопротеидных комплексов и может быть труднодоступной для антисмысловых РНК, в связи с чем потребуется разработка дополнительных условий для повышения эффективности рибозимов, включенных в состав антисмысловых РНК.