Фенотип организма определяется совокупностью многих признаков, за развитие которых отвечают разные гены, принадлежащие одной, занимая в ней различные локусы, или разным группам сцепления (хромосомам). При этом обнаруживаются признаки, которые наследуются как независимо друг относительно друга, так и совместно (сцеплено).
Закон независимого наследования признаков сформулирован Г. Менделем по результатам наблюдений за наследованием одновременно двух признаков - цвета и формы горошин. Названный закон утверждает, что различные признаки, контролируемые неаллельными генами

Рис. 4.33. Наследование, сцепленное с полом (окраска глаз у дрозофилы):
I - сочетание половых хромосом в кариотипе представителей разного пола;
II - гомогаметный пол образует один тип гамет, гетерогаметный - два; III - представители гомогаметного пола получают хромосомы Х от обоих родителей, представители гетерогаметного пола получают единственную хромосому Х от гомогаметного родителя, а хромосому Y - от гетерогаметного родителя. Окрашены отцовские хромосомы

Рис. 4.34. Обоснование особенностей Х-сцепленного и У-сцепленного типов наследования поведением половых хромосом при образовании гамет особями гомо- и гетерогаметного пола и при оплодотворении: I - сочетание половых хромосом в кариотипах представителей разных полов; II - гомогаметный пол образует один тип гамет, гетерогаметный два; III - представители гомогаметно-го пола получают хромосомы от обоих родителей; представители гетерогамет-ного пола получают хромосому Х от гомогаметного родителя, а хромосому У от гетерогаметного родителя; это справедливо для генов, расположенных в негомологичных локусах хромосом Х и У; окрашены отцовские хромосомы
(наследственными задатками), передаются от родителей потомству независимо друг от друга и обнаруживаются в фенотипах потомков во всех возможных сочетаниях (рис. 4.35).
Очевидно, что этому закону должны подчиняться признаки, развитие которых контролируют неаллельные гены, находящиеся в разных (негомологичных) хромосомах. В таком случае независимый характер наследования двух признаков или более объясняется поведением негомологичных хромосом в мейозе. Названные хромосомы образуют со своими гомологами в первом делении мейоза пары (биваленты). В анафазе I мейоза гомологи каждой пары расходятся к полюсам делящейся клетки независимо от гомологов других пар. В результате гаплоидные хромосомные наборы будущих гамет на полюсах представлены случайными сочетаниями отцовских и материнских хромосом. Следовательно, различные гаметы содержат разные комбинации отцовских и материнских аллелей неаллельных генов.
Разнообразие вариантов образуемых гамет зависит от степени ге-терозиготности организма и описывается формулой 2n, где n - число

Рис. 4.35. Независимое наследование двух признаков (цвет и форма горошин)
локусов в гетерозиготном состоянии. В связи с этим дигетерозиготные гибриды первого поколения (F1), рождаемые в скрещиваниях гомозиготных, по доминантным (один родитель - АВ) и рецессивным (второй родитель - ав) аллелям анализируемых генов, проявляя в силу гетеро-зиготности и свойства доминантности-рецессивности аллелей (АаВв) фенотипическое единообразие по обоим наблюдаемым признакам, образуют четыре варианта гамет, причем с равной вероятностью каждо-
го из вариантов. При скрещивании гибридов F1 между собой благодаря всем возможным комбинациям аллелей, присутствующих в гаметах указанных четырех вариантов, в случае независимого наследования двух признаков (дигибридное скрещивание) среди гибридов второго поколения (F2) обнаруживается четыре фенотипические группы (в сравнении с F1 происходит расщепление по фенотипу) потомков в отношении 9:3:3:1. Анализ потомства F2 отдельно по каждой из двух наблюдаемых пар альтернативных признаков - цвет (зеленый или желтый) и форма (гладкая или морщинистая) горошин - выявляет наличие по каждой паре двух фенотипических групп (расщепление по фенотипу) потомков в отношении 3:1 (моногибридное скрещивание).
В опытах Г. Менделя наследственная конституция гибридов F1 (АаВв) устанавливалась путем анализа фенотипов потомства F2, получаемого при самоопылении растений-родителей (F1). Эта же задача решается, если применяется так называемое анализирующее скрещивание. Оно заключается в скрещивании организма, генотип которого необходимо установить, с организмом-гомозиготой по рецессивному(ым) аллелю(ям) соответствующего(их) гена(ов) - рис. 4.36 (моногибридное анализирующее скрещивание) и рис. 4.37 (дигибридное анализирующее скрещивание). Так как гомозиготные родители образуют один тип гамет: аа-а (моногибридное скрещивание), ааЪЪ-аЪ (дигибридное скрещивание), ааЪЪсс-аЪс (тригибридное скрещивание) и т.д., - при анализирующем скрещивании количество разных фенотипов потомков зависит от числа типов гамет организма с доминантным фенотипом. Если последний гомозиготен по анализируемым генам, то он тоже образует только один тип гамет, и потомство в анализирующем скрещивании отличается фенотипическим единообразием, так как все потомки имеют доминантный фенотип (см. рис. 4.35, I). Если организм, генотип которого необходимо установить, гетерозиготен по одному гену, он образует два типа гамет, в силу чего при анализирующем скрещивании рождают-

Рис. 4.36. Анализирующее моногибридное скрещивание. Объяснения в тексте
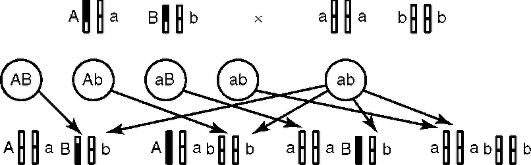
Рис. 4.37. Анализирующее дигибридное скрещивание. Объяснения в тексте
ся потомки двух разных фенотипов - один с доминантным и второй с рецессивным признаком (см. рис. 4.35, II). Дигетерозиготный организм дает при анализирующем скрещивании потомство с четырьмя разными фенотипами (см. рис. 4.37).
Признаки, контролируемые генами, находящимися в одной хромосоме, могут демонстрировать как независимый, если соответствующие локусы расположены относительно далеко - 50 морганид (сантиморга-нид) и более, так и сцепленный тип наследования. В такой ситуации признаки передаются потомству всегда или в определенном проценте случаев совместно. Подозрение в том, что феномен сцепленного наследования существует, возникло, когда было обнаружено, что результаты анализирующего скрещивания гибридов F1 у дрозофил иногда отличались от ожидаемых, если исходить из исключительно независимого соотносительного наследования нескольких признаков (Т.Г. Морган). Конкретно у потомков в таких скрещиваниях вместо ожидаемого свободного комбинирования фенотипических признаков, контролируемых аллельными парами разных генов, наблюдали тенденцию к наследованию преимущественно родительских сочетаний признаков. Как уже отмечалось, в основе сцепленного соотносительного наследования признаков лежит расположение соответствующих генов в одной хромосоме. Именно это обстоятельство привело к тому, что каждая хромосома стала рассматриваться как отдельная группа сцепления. На рисунке 4.38 представлены результаты наследования признаков окраски тела и формы (длины) крыльев у дрозофилы, а также цитологическое обоснование этих результатов. При анализирующем скрещивании самцов из F1 появлялось всего два фенотипических вида потомков, сходных с родительскими формами по сочетанию вариантов анализируемых признаков (серая окраска тела и нормальные крылья; черная окраска тела и короткие крылья) в соотношении 1:1. Это указывает на образо-

Рис. 4.38. Сцепленное наследование признаков (окраска тела и длина крыльев) у дрозофилы: I - скрещивание чистых линий; II и III - анализирующие скрещивания, соответственно самцов и самок из F1
вание (с одинаковой вероятностью) самцами F1 всего двух типов гамет, в которые попадают исходные (родительские) сочетания аллелей генов, контролирующих развитие названных признаков - окраска тела (В и b) и форма или длина крыльев (V и v). При анализирующем скрещивании самок F1 обнаруживалось четыре фенотипических варианта потомков, практически со всеми возможными сочетаниями признаков. При этом потомки с родительским сочетанием признаков составили 83%. У 17% потомков - кроссоверное потомство - обнаруживались иные комбинации признаков (серая окраска тела и короткие крылья; черная окраска тела и нормальные крылья). Видно, что в указанных скрещиваниях проявляется склонность к сцепленному наследованию либо доминантных, либо рецессивных признаков (83%). Частичное (17%) нарушение сцепления наследования признаков потомками объясняется процессом кроссинговера - обменом гомологичными участками между гомологичными хромосомами в профазе I мейоза.
Сцепленное соотносительное наследование двух признаков или более не следует путать с наследованием, сцепленным с полом. Напомним, что наследование, сцепленное с полом, выделяется на основании одного критерия - локализации соответствующего гена в половой хромосоме (Х или У).
Анализ соотносительного наследования сочетаний других признаков, прежде всего у плодовых мух дрозофил, показал, что процент кроссоверного потомства для каждой пары признаков всегда один и тот же, но различается для разных пар. Это послужило основанием для заключения о том, что гены («наследственные задатки», по Г. Менделю) располагаются в хромосомах в линейном порядке, причем разные (негомологичные) хромосомы представляют собой группы сцепления определенных генов. Напротив, гомологичные хромосомы - это группы сцепления одних и тех же генов, которые, однако, в гомологах могут быть представлены разными аллелями. В профазе I мейоза гомологи каждой пары конъюгируют, т. е. сближаются с точным противостоянием гомологичных локусов (другими словами, аллелей соответствующего гена). Затем благодаря кроссинговеру хромосомы в парах гомологов могут обменяться гомологичными участками. Если обмениваемые участки гомологичных хромосом представлены разными аллелями гена, занимающего локус, обмен приводит к изменению аллельного состава каждой из гомологичных хромосом (один из факторов генотипической комбинативной изменчивости). Следовательно, потомок, получив в результате акта оплодотворения хромосому с измененным аллельным на-

Рис. 4.39. Схема, поясняющая низкий процент кроссоверных гамет для двух генов. Плюсом обозначены клетки-предшественницы гамет, в которых на участке между двумя избранными локусами произошел кроссинговер; красным цветом обозначены кроссоверные гаметы
бором, будет отличаться от родителя определенными фенотипическими особенностями.
Частота, с которой происходит обмен на участке хромосомы между двумя конкретными генами, зависит от расстояния между ними (правило Т. Моргана). Другими словами, при увеличении расстояния между генами одной хромосомы (группы сцепления) вероятность кроссинго-вера между ними растет, однако даже в случае осуществления обмена между генами данной пары во всех клетках-предшественницах половых клеток процент кроссоверных гамет не превышает 50. Это происходит потому, что в акте кроссинговера участвуют две хроматиды из четырех, имеющихся в каждом биваленте (рис. 4.39). С увеличением расстояния между генами в группах сцепления растет вероятность того, что на соответствующем участке одновременно произойдет несколько кроссин-говеров. Так как каждый второй перекрест (кроссинговер) приводит к восстановлению в хромосоме прежнего сочетания аллелей, то с ростом расстояния между генами количество кроссоверных гамет может не увеличиваться, а уменьшаться. Следовательно, процент кроссоверных гамет представляет собой адекватный показатель расстояния между двумя генами, только если они находятся на достаточно близком расстоянии, когда исключается вероятность второго кроссинговера.
Различают неполное (частичное) и полное сцепление. Полное сцепление (фактически отсутствие кроссинговера) может быть видовой характеристикой гаметогенеза у представителей одного из полов, например у самцов дрозофилы.
Применение анализирующего скрещивания дает возможность выяснить не только генный состав отдельных групп сцепления (хромосом), но также установить расстояние между генами.
38. Виды взаимодействия аллельных генов. Примеры у человека.
Взаимодействие аллельных генов.
Постоянно возникающие разнообразные изменения структуры генов обусловливают явление множественного аллелизма, поэтому взаимодействующие при оплодотворении гаметы часто несут в своих геномах разные аллели одного и того же гена. В связи с этим генотип нового организма является гетерозиготным по многим локусам, т.е. его аллельные гены, расположенные в соответствующих участках гомологичных хромосом и пришедшие от обоих родителей, представлены разными аллелями— А и А', В и В' С и С' Если аллельные гены представлены одинаковыми аллелями, т.е. находятся в гомозиготном состоянии, то развивается соответствующий данному аллелю вариант признака. В случае гетерозиготности развитие данного признака будет зависеть от взаимодействия аллельных генов.
Доминирование
Называется тип взаимодействия аллейных генов, при котором один (доминантный) аллейный ген у гибридов первого поколения полностью подавляет проявление другого (рецессивного) аллейного гена.
Примером доминирования одного из аллелей в гетерозиготном генотипе может
служить определение групповой принадлежности крови у человека по системе АВ0.
Неполное доминирование
Наблюдается, когда фенотип гетерозигот BB' отличается от фенотипа гомозигот по обоим аллелям (BB или B'B') Промежуточным проявлением признака. Это объясняется тем, что аллель, способный сформировать нормальный признак, находясь в двойной дозе у гомозиготы BB, проявляется сильнее, чем в единственной дозе у гетерозиготы BB'. Указанные генотипы отличаются экспрессивностью, т.е. степенью выраженности признака.
Демонстрацией такого типа взаимодействия генов могут быть многочисленные наследственные заболевания у человека, проявляющиеся клинически у гетерозигот по мутантным аллелям, а у гомозигот заканчивающиеся смертью. Иногда гетерозиготы имеют почти нормальный фенотип, а гомозиготы характеризуются пониженной жизнеспособностью. Так, гомозиготы по аллелю серповидноклеточности эритроцитов в связи с развитием у них тяжелой формы анемии и других фенотипических проявлений
обычно не переживают детский возраст. Напротив, гетерозиготы—это как правило нормальные люди.
Кодоминирование
Представляет собой такой тип взаимодействия аллельных генов, при котором каждый из аллелей проявляет свое действие. В результате этого формируется некий промежуточный вариант признака, новый по сравнению с вариантами, определяемыми каждым аллелем самостоятельно. Примером может служить формирование IV, или АВ-группы, крови у человека, гетерозиготного по
аллелям IA иIB, которые по отдельности детерминируют образование II и III групп крови.