Система регуляции экспрессии генов у эукариот связана с особенностями функционирования эукариотического генома. Хотя и упрокариот, и у эукариот функционируют системы регуляторных белков, наличие ядра и нуклеосомная организация хроматина эукариот дают намного больше возможностей для регуляции. Ядро явилось важнейшим эволюционным приобретением эукариот. Благодаря ядерной мембране, разделяются зоны транскрипции и трансляции, что позволяет осуществлять сложную и многообразную регуляцию экспрессии генов. Такая регуляция происходит на всех этапах.
Регуляция на уровне транскрипции. Основным уровнем регуляции экспрессии уэукариот является регуляция на уровне транскрипции. Варианты такой регуляции весьма разнообразны.
Наиболее универсальным методом регуляции транскрипции служит конденсация-деконденсация хроматина. Хорошо известно, что при активации генетического материала он деконденсируется. С другой стороны, далеко не весь эухроматин транскрибируется. Поэтому имеются и другие пути контроля транскрипции.
Дляэукариот характерна дифференциальная экспрессия генов в разных клетках организма, поэтому многоклеточные организмы имеют разнообразные дифференцированные клетки. Хотя во всех клетках содержатся одинаковые гены, экспрессируются они не одинаково. Основное направление регуляции клеточной дифференцировки осуществляется при помощи тканеспецифических транскрипционных факторов. Они представляют собой белки регуляторных генов, контролирующие дифференциальную активность других генов.
Иным направлением является использование альтернативных промоторов. В разных типах клеток с одного гена могут быть образованы разные белки. Так, ген белка дистрофина содержит 7 промоторов, которые считывают информацию, начиная с разных экзонов.
Большое значение в регуляции экспрессии генов уэукариот придается фактору метилирования ДНК внутри регуляторных областей. Метилированию подвергается цитозин в составе ЦГ-динуклеотида, что обычно приводит к инактивации гена. Деметилирование ДНК восстанавливает активность. Этот важный процесс регулируют ферменты метилтрансферазы.
Частным случаем регуляции на уровне транскрипции является гормональная регуляция, при которой гены «включаются» в ответ на внешний сигнал. Сигнал запускает экспрессию только тех генов, которые имеют специфические последовательности ДНК в своих регуляторных областях.
Весьма загадочным способом регуляции является геномный импринтинг – дифференциальное проявление отцовских и материнских генов в организме. Геномыбудто бы «запоминают» свое происхождение. Механизм этого явления не установлен.
С данным явлением схожа инактивация Х-хромосомы в соматических клетках женского организма млекопитающих. Правда, одна из двух Х-хромосом в клетках плода инактивируется случайным образом, поэтому в разных клетках женского организма экспрессируются как отцовские, так и материнские Х-хромосомы.
На процессы «включения» и «выключения» генов, их конденсации и деконденсации ощутимое влияние оказывают различные хромосомные перестройки, МГЭ, изменяющие эффект положения гена.
Дляэукариот не характерна регуляция по типу оперона. Большинство м-РНК эукариот – моноцистронные. Более того, гены, контролирующие один метаболический путь, у эукариот часто разбросаны по всему геному. Но в этом правиле есть исключение. Гены р-РНК и митохондрий транскрибируются на единую транскрипционную единицу, которая разрезается после процессинга. Считается, что такой способ позволяет унифицировать регуляцию схожих генов.
Регуляция на уровне процессинга. Уэукариот транскрипция гена еще не означает его проявления в фенотипе. Молекулы РНК, синтезированные в ходе транскрипции, у эукариот подвергаются существенным модификациям. Совокупность таких преобразований и составляет сущность процессинга.
На уровне процессинга в первую очередь необходимо отметить механизм альтернативного сплайсинга, позволяющий изменять порядок сшивки экзонов. Таким образом, на основе одной и той же нуклеотидной последовательности одного гена формируются разные белки, состоящие из разных сочетаний одних и тех же аминокислотных блоков.
Феномен альтернативного сплайсинга изменил наши представления о природе гена. Его распространенность оказалась значительно выше, а роль в эволюции – важнее, чем предполагалось первоначально. Альтернативный сплайсинг характерен для более чем трети генов человека. Так, на гене белка тропонина, содержащего 18 экзонов, за счет альтернативного сплайсинга может образовываться 64 различных белковых продукта. В среднем один ген человека кодирует около трех белков (Тарантул В. З., 2003).
Другим интересным способом регуляции на уровне процессинга является тканеспецифическое редактирование РНК. Оно обнаружено у микроорганизмов, грибов, млекопитающих и проявляется заменой отдельных нуклеотидов в молекуле РНК при помощи специального ферментного комплекса. Если в случае замены вместо смыслового кодона образуется стоп-кодон, то в новой полипептидной цепи будут отсутствовать все аминокислоты, идущие после него. Получается белок с совершенно новыми свойствами.
В ооцитах некоторых животных происходит накопление и-РНК, у которых не закончен процессинг. Такие РНК не транслируются. Окончание процессинга и последующая экспрессия наступают только после оплодотворения.
Примерно половина гя-РНК полностью распадается в ядре, не выходя за его пределы. Возможно распаду подвержены такие транскрипты, которые не способны превратиться в зрелую и-РНК.
Регуляция на уровне трансляции. Механизмы регуляции экспрессии на уровне трансляции изучены недостаточно полно. Избирательная трансляция м-РНК осуществляется отбором определенных м-РНК путем блокировки доступа к рибосомам.
В случае избирательной стабилизации определенных типов м-РНК в цитоплазме, они не подвергаются распаду после трансляции. Наглядным примером дифференциальной стабильности м-РНК может служить синтез белков глобинов на стабильных м-РНК безъядерных ретикулоцитов млекопитающих. Мутации, блокирующие полиаденилирование, наоборот, приводят к образованию очень нестабильных транскриптов. Обнаружены и другие механизмы, регулирующие стабильность м-РНК.
Регуляция на уровне посттрансляционной модификации белка. Посттрансляционная модификация полипептида и превращение его в функционально активную молекулу белка завершает процесс реализации генетической информации. Она представляет собой различные модификации определенных аминокислот (фосфорилирование, ацетилирование), удаление некоторых из них, и на этой основе формирование вторичной, третичной, четвертичной структуры белка. На посттрансляционном уровне также возможна регуляция экспрессии. Широко распространен механизм регуляции активности ферментов, основанный на присоединении молекул-эффекторов, в роли которых часто выступают конечные продукты биосинтеза.
Основная масса генов, активно функционирующих в большинстве клеток организма на протяжении онтогенеза, — это гены, которые обеспечивают синтез белков общего назначения (белки рибосом, гистоны, тубулины и т.д.), тРНК и рРНК. Транскрибирование этих генов обеспечивается соединением РНК-полимеразы с их промоторами и, видимо, не подчиняется каким-либо другим регулирующим воздействиям. Такие гены называют конститутивными. Другая группа генов, детерминирующих синтез специфических продуктов, в своем функционировании зависит от различных регулирующих факторов, ее называют регулируемыми генами (рис. 3.84). Их активное функционирование, скорость и продолжительность транскрипции регулируются путем стимуляции или запрещения соединения РНК-полимеразы с промоторной областью гена.
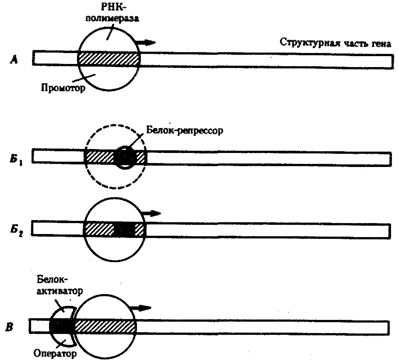
Рис. 3.84. Схема конститутивных и регулируемых генов:
А — конститутивный ген; Б, В — регулируемые гены; Б1Б2 —связывание РНК-полимеразы с промотором возможно лишь в отсутствие белка-репрессора, который специфически соединяется с оператором, частично или полностью перекрывающим промоторную последовательность; В —связывание РНК-полимеразы с промотором облегчается белком-активатором (апоиндуктором), который специфически узнает область оператора, расположенную перед промотором; стрелкой обозначено направление транскрипции
46. Особенности человека как объекта генетического анализа. Методы изучения генетики человека: генеалогический, биохимический, цитогенетический, дерматоглифический,ДНК-диагностики, генетики соматических клеток, популяционно статистический, близнецовый.
Для применения гибридологического метода генетического анализа, человек как вид обладает целым рядом особенностей, не позволяющих применять этот метод для изучения его наследственности и изменчивости. Во-первых, у человека не может быть произведено искусственного направленного скрещивания в интересах исследователя. Во-вторых, низкая плодовитость делает невозможным применение статистического подхода при оценке немногочисленного потомства одной пары родителей. В-третьих, редкая смена поколений, происходящая в среднем через 25 лет, при значительной продолжительности жизни дает возможность одному исследователю наблюдать не более 3—4 последовательных поколений. Изучение генетики человека затрудняется наличием в его геноме большого числа групп сцепления генов (23 у женщин и 24 у мужчин), а также высокой степенью фенотипического полиморфизма, связанного с влиянием среды.
К методам, широко используемым при изучении генетики человека, относятся генеалогический, популяционно-статистический, близнецовый, метод дерматоглифики, цитогенетический, биохимический, методы генетики соматических клеток.
Близнецовый метод. Этот метод заключается в изучении закономерностей наследования признаков в парах одно- и двуяйцевых близнецов.
Суть метода заключается в сравнении проявления признака в разных группах близнецов при учете сходства или различия их генотипов. Монозиготные близнецы, развивающиеся из одной оплодотворенной яйцеклетки, генетически идентичны, так как имеют 100% общих генов. Поэтому среди монозиготных близнецов наблюдается высокий процент конкордантных пар, в которых признак развивается у обоих близнецов. Сравнение монозиготных близнецов, воспитывающихся в разных условиях постэмбрионального периода, позволяет выявить признаки, в формировании которых существенная роль принадлежит факторам среды. По этим признакам между близнецами наблюдается дискордантность, т.е. различия. Напротив, сохранение сходства между близнецами, несмотря на различия условий их существования, свидетельствует о наследственной обусловленности признака.
Применение: изучение наследственности и изменчивости у человека для определения соотносительной роли наследственности и среды в формировании различных признаков, как нормальных, так и патологических, оценить эффективность действия на организм некоторых внешних факторов (лекарственных препаратов, обучения, воспитания).
Популяционно-статистический метод
С помощью популяционно-статистического метода изучают наследственные признаки в больших группах населения, в одном или нескольких поколениях. Существенным моментом при использовании этого метода является статистическая обработка получаемых данных. Этим методом можно рассчитать частоту встречаемости в популяции различных аллелей гена и разных генотипов по этим аллелям, выяснить распространение в ней различных наследственных признаков, в том числе заболеваний. Он позволяет изучать мутационный процесс, роль наследственности и среды в формировании фенотипического полиморфизма человека по нормальным признакам, а также в возникновении болезней, особенно с наследственной предрасположенностью. Этот метод используют и для выяснения значения генетических факторов в антропогенезе, в частности в расообразовании.
При статистической обработке материала, получаемого при обследовании группы населения по интересующему исследователя признаку, основой для выяснения генетической структуры популяции является закон генетического равновесия Харди — Вайнберга. Он отражает закономерность, в соответствии с которой при определенных условиях соотношение аллелей генов и генотипов в генофонде популяции сохраняется неизменным в ряду поколений этой популяции.
Математическим выражением закона Харди — Вайнберга служит формула (рА. + qa)2, где р и q — частоты встречаемости аллелей А и а соответствующего гена. Раскрытие этой формулы дает возможность рассчитать частоту встречаемости людей с разным генотипом и в первую очередь гетерозигот — носителей скрытого рецессивного аллеля: p2AA + 2pqAa + q2аа.
ДНК-диагностика - это совокупность методов и технологий, которые позволяют выявлять повреждения в определенном гене человека, который либо является причиной различных заболеваний, либо может к ним привести.
Для ДНК-диагностики подходят следующие виды биологических материалов: кровь, букальный эпителий, слюна, сперма, ворсины хориона и амниотическая жидкость, образцы различных органов и тканей, содержащих целые клетки с неразрушенными ядрами. Можно использовать пятна крови, нанесенные на пористую поверхность, а также трупный и эксгумированный материал. Однако, чаще всего источником ДНК служат клетки крови и плодный материал.
ДНК-диагностику используют в следующих случаях:
· Медико-генетическая консультация при наследственных заболеваниях. Проводится в случаях наличия в семье ребенка или близкого родственника с наследственным заболеванием. Диагностикапроводится с целью избежать рождение больного ребенка.
· Установление биологического родства. Экспертиза позволяет со 100 %-ной вероятностью исключить отцовство (материнство) или с вероятностью не ниже 99,99 % установить наличие биологического родства. В общем случае срок проведения анализа 2 недели с момента поступления образцов в лабораторию. Такая диагностика возможна только по постановлению следственных и судебных органов или в независимых лабораториях.
Диагностика к предрасположенности к некоторым заболеваниям, таким как атеросклероз, ишемическая болезнь сердца, эссенциальная гипертензия, остеопороз, сахарный диабет, бронхиальная астма, рак и пр. Генетическое тестирование не позволяет установить диагноз, а дает возможность выявить наличие тенденции будущих болезней и, исходя из современного врачебного опыта, наметить пути их ранней профилактики.
· Диагностика некоторых форм бесплодия и невынашивания беременности
· Определение ДНК возбудителя при различных бактериальных и вирусных инфекциях
Возбудители практически всех инфекционных болезней вирусы и бактерии также обладают ДНК или РНК, что позволяет проводить их диагностику аналогичными методами. Использование этих методов для определения возбудителей урогенитальных инфекций, многие из которых часто протекают бессимптомно, но приводят к неблагоприятным последствиям, помогло поднять их диагностику на принципиально иной уровень.
Для генетических исследований человек является неудобным объектом, так как у человека: невозможно экспериментальное скрещивание; большое количество хромосом; поздно наступает половая зрелость; малое число потомков в каждой семье; невозможно уравнивание условий жизни для потомства.
В генетике человека используется ряд методов исследования.
Генеалогический метод
Использование этого метода возможно в том случае, когда известны прямые родственники — предки обладателя наследственного признака (пробанда) по материнской и отцовской линиям в ряду поколений или потомки пробанда также в нескольких поколениях. При составлении родословных в генетике используется определенная система обозначений. После составления родословной проводится ее анализ с целью установления характера наследования изучаемого признака.

Условные обозначения, принятые при составлении родословных:
1 — мужчина; 2 — женщина; 3 — пол не выяснен; 4 — обладатель изучаемого признака; 5 — гетерозиготный носитель изучаемого рецессивного гена; 6 — брак; 7 — брак мужчины с двумя женщинами; 8 — родственный брак; 9 — родители, дети и порядок их рождения; 10 — дизиготные близнецы; 11 — монозиготные близнецы.
Благодаря генеалогическому методу были определены типы наследования многих признаков у человека. Так, по аутосомно-доминантному типу наследуются полидактилия (увеличенное количество пальцев), возможность свертывать язык в трубочку, брахидактилия (короткопалость, обусловленная отсутствием двух фаланг на пальцах), веснушки, раннее облысение, сросшиеся пальцы, заячья губа, волчья пасть, катаракта глаз, хрупкость костей и многие другие. Альбинизм, рыжие волосы, подверженность полиомиелиту, сахарный диабет, врожденная глухота и другие признаки наследуются как аутосомно-рецессивные.

Доминантный признак — способность свертывать язык в трубочку (1) и его рецессивный аллель — отсутствие этой способности (2).
3 — родословная по полидактилии (аутосомно-доминантное наследование).
Целый ряд признаков наследуется сцепленно с полом: Х -сцепленное наследование — гемофилия, дальтонизм; Y -сцепленное — гипертрихоз края ушной раковины, перепончатость пальцев ног. Имеется ряд генов, локализованных в гомологичных участках Х - и Y -хромосом, например общая цветовая слепота.
Использование генеалогического метода показало, что при родственном браке, по сравнению с неродственным, значительно возрастает вероятность появления уродств, мертворождений, ранней смертности в потомстве. В родственных браках рецессивные гены чаще переходят в гомозиготное состояние, в результате развиваются те или иные аномалии. Примером этого является наследование гемофилии в царских домах Европы.

Наследование гемофилии в царских домах Европы:
 — гемофилик;
— гемофилик;  — женщина-носитель.
— женщина-носитель.
Цитогенетический метод
Основан на изучении хромосом человека в норме и при патологии. В норме кариотип человека включает 46 хромосом — 22 пары аутосом и две половые хромосомы. Использование данного метода позволило выявить группу болезней, связанных либо с изменением числа хромосом, либо с изменениями их структуры. Такие болезни получили название хромосомных.
Материалом для кариотипического анализа чаще всего являются лимфоциты крови. Кровь берется у взрослых из вены, у новорожденных — из пальца, мочки уха или пятки. Лимфоциты культивируются в особой питательной среде, в состав которой, в частности, добавлены вещества, «заставляющие» лимфоциты интенсивно делиться митозом. Через некоторое время в культуру клеток добавляют колхицин. Колхицин останавливает митоз на уровне метафазы. Именно во время метафазы хромосомы являются наиболее конденсированными. Далее клетки переносятся на предметные стекла, сушатся и окрашиваются различными красителями. Окраска может быть а) рутинной (хромосомы окрашиваются равномерно), б) дифференциальной (хромосомы приобретают поперечную исчерченность, причем каждая хромосома имеет индивидуальный рисунок). Рутинная окраска позволяет выявить геномные мутации, определить групповую принадлежность хромосомы, узнать, в какой группе изменилось число хромосом. Дифференциальная окраска позволяет выявить хромосомные мутации, определить хромосому до номера, выяснить вид хромосомной мутации.
В тех случаях, когда необходимо провести кариотипический анализ плода, для культивирования берутся клетки амниотической (околоплодной) жидкости — смесь фибробластоподобных и эпителиальных клеток.
К числу хромосомных заболеваний относятся: синдром Клайнфельтера, синдром Тернера-Шерешевского, синдром Дауна, синдром Патау, синдром Эдвардса и другие.
Больные с синдромом Клайнфельтера (47, ХХY) всегда мужчины. Они характеризуются недоразвитием половых желез, дегенерацией семенных канальцев, часто умственной отсталостью, высоким ростом (за счет непропорционально длинных ног).
Синдром Тернера-Шерешевского (45, Х0) наблюдается у женщин. Он проявляется в замедлении полового созревания, недоразвитии половых желез, аменорее (отсутствии менструаций), бесплодии. Женщины с синдромом Тернера-Шерешевского имеют малый рост, тело диспропорционально — более развита верхняя часть тела, плечи широкие, таз узкий — нижние конечности укорочены, шея короткая со складками, «монголоидный» разрез глаз и ряд других признаков.
Синдром Дауна — одна из самых часто встречающихся хромосомных болезней. Она развивается в результате трисомии по 21 хромосоме (47; 21, 21, 21). Болезнь легко диагностируется, так как имеет ряд характерных признаков: укороченные конечности, маленький череп, плоское, широкое переносье, узкие глазные щели с косым разрезом, наличие складки верхнего века, психическая отсталость. Часто наблюдаются и нарушения строения внутренних органов.
Хромосомные болезни возникают и в результате изменения самих хромосом. Так, делеция р -плеча аутосомы №5 приводит к развитию синдрома «крик кошки». У детей с этим синдромом нарушается строение гортани, и они в раннем детстве имеют своеобразный «мяукающий» тембр голоса. Кроме того, наблюдается отсталость психомоторного развития и слабоумие.
Чаще всего хромосомные болезни являются результатом мутаций, произошедших в половых клетках одного из родителей.
Биохимический метод
Позволяет обнаружить нарушения в обмене веществ, вызванные изменением генов и, как следствие, изменением активности различных ферментов. Наследственные болезни обмена веществ подразделяются на болезни углеводного обмена (сахарный диабет), обмена аминокислот, липидов, минералов и др.
Фенилкетонурия относится к болезням аминокислотного обмена. Блокируется превращение незаменимой аминокислоты фенилаланин в тирозин, при этом фенилаланин превращается в фенилпировиноградную кислоту, которая выводится с мочой. Заболевание приводит к быстрому развитию слабоумия у детей. Ранняя диагностика и диета позволяют приостановить развитие заболевания.
Метод дерматоглифики
Дерматоглифка – это изучение рельефа кожи на пальцах, ладонях и подошвенных поверхностях стоп, который образован эпидермальными выступами – гребнями, которые образуют сложные узоры.
Ф. Гальтон предложил предложил классификацию этих узоров, позволившую использовать этот метод для идентификации личности в криминалистике.
Разделы дерматоглифики:
•дактилоскопия – изучение узоров на подушечках пальцев
•пальмоскопия – изучение рисунка на ладонях
•плантоскопия – изучение дерматоглифики подошвенной поверхности стопы
Дактилоскопия. Гребни на коже пальцев рук соответствуют сосочкам дермы, поэтому их называют также папиллярными линиями, рельеф этих выступов повторяет пласт эпидермиса. Межсосочковые углубления образуют бороздки. Закладка узоров происходит между 10 и 19 неделями внутриутробного развития; у 20 недельных плодов уже хорошо различимы формы узоров. Формирование папиллярного рельефа зависит от характера ветвления нервных волокон. Полное формирование деталей строения тактильных узоров отмечается к шести месяцам, после чего они остаются неизменными до конца жизни. Дерматоглифические исследования имеют важное значение в определении зиготности близнецов, в диагностике некоторых наследственных заболеваний, в судебной медицине, в криминалистике для идентификации личности.
Пальмоскопия. Ладонный рельеф очень сложный, в нем выделяют ряд полей, подушечек и ладонных линий. У правшей более сложные узоры встречаются на правой руке, у левшей – на левой. Индивидуальные особенности кожных узоров наследственно обусловлены. Это доказано многими генетическими исследованиями, в частности, на монозиготных близнецах.
Обширные исследования по изучению особенностей дерматоглифики проведены у нас в стране Т.Д. Гладковой (1996), а по наследственной обусловленности кожных узоров – И.С. Гусевой (1970, 1980).на основании этих работ был сделан вывод, что количественные показатели рельефа гребневой кожи программируются полигенной системой, включающей небольшое число аддитивно действующих генов. гены гребневой кожи проявляют свой морфогенетический эффект, влияя на степень ветвления нервного волокна, и фенотипически определяют гребневую плотность. На формирование дерматоглифических узоров могут оказывать влияние некоторые повреждающие факторы на ранних стадиях эмбрионального развития.