Штаммы типа I Т. гондий, которые остро вирулентными у мышей (Сибли и Бутройда, 1992), содержит две изоформы NTPase (Бермудес и др.,

200 БИОХИМИЯ И ОБМЕН токсоплазма
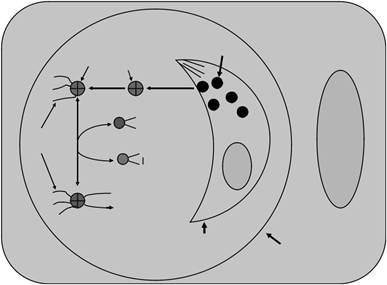
клетка-хозяин
NTPase DG
(В состоянии покоя)
SH
SH
| ИВН | N |
S
S N
NTP, NDP
NDP, NMP
NTPase
(Активный)
тахизоит PVM
Рисунок 8.7 Поведение NTPase в тахизоит-инфицированной клетки. NTPase секретируется из плотной гранулы тахизоит в паразитофорное вакуолярном пространство и ассоциированных к intravacuolar сети. Затем активность NTPase может регулироваться изменением окислительно-восстановительного в своей молекуле, пострадавшими от дитиольному соединения или неизвестной дитиольному-дисульфида оксидоредуктазы в пределах паразитофорной вакуоли. Сокращения, используемые в следующем: DG, плотные гранулы; ПВМ, паразитофорная вакуоль мембрана; IVN, intravacuolar сеть; N, ядро.
1994; Асаи и др., 1995). Одна из изоформ (NTPase-я в Асаи и др., 1995;. NTP3 в Bermudes и др, 1994) преимущественно гидролизует трифосфат нуклеозидов, а другой изоформы (NTPase-II в Асаи и др., 1995; NTP1 в Бермудес и др., 1994) гидролизует три- и дифосфат нуклеозиды на две приблизительно равные ставки. Изоформы NTPase-я, кажется, присутствуют только в I вирулентных штаммов типа, в то время как NTPase-II универсально присутствует во всех штаммах Т. гондий (Асаи и др., 1995). Хотя присутствие NTPase-I, кажется, один из основных факторов вирулентности у мышей, нет прямых доказательств роли в вирулентности не было доказано. Neospora сатпит, который авирулентная у мышей, имеет только тип фермента NTPase-I, и ни один тип фермента NTPase-II, не обнаружено (Асаи и др., 1998).
Полный кДНК для NTPase-I и II NTPase-кодирований предсказали открытые рамки считывания одинакового размера, которые отличаются в 16 из 628 аминокислот между два изоформом (Асаи и др., 1995). Молекулярная масса
нативные NTPases примерно 260 кДа, состоящий из четырех идентичных субъединиц с предсказанной молекулярной массой 67 кДа (Асаи и др., 1995). Обе изоформы NTPase содержат N-концевой гидрофобный сигнальный пептид (25 аминокислот), который отсутствует в нативном NTPase очищенного от тахи- zoites (Асаи и др., 1995). Предполагается, что этот сигнальный пептид служит сигналом для транспортировки NTPase к плотной грануле. Другие плотные гранулы-белки имеют сходные N-концевые гидрофобные сигнальные пептиды (Cesbron-Delauw и др., 1996).
Как уже упоминалось выше, основное различие между этими NTPase изоформ заключается в их способности к гидролизу нуклеозид трифосфат против дифосфат субстратов. В то время как NTPase-II гидролизует АТФ в АДФ и АДФ до АМФ при почти той же скоростью, как нативный и рекомбинантный NTPase-ли гидролизуют АДФ до АМФ с гораздо меньшей скоростью - менее 1 процента от скорости для АТФ (Асаи и др. 1995, 1998). Это говорит о том, что Т. гондий NTPase должен
| РЕКОМЕНДАЦИИ |
классифицироваться как NTP diphosphohydrolase (апиразы, EC 3.6.1.5), новый ген семейства АТФаз E-типа. Т. гондий NTPase имеет гомологию с апираза, и антитело против Т. гондий NTPase признает apyrases картофеля и Schistosoma mansoni (Васконселос и др., 1996), и Trypanosoma Тгурапозота (Fietto и др., 2004). Обильная ДТТЫ-активированная NTPase в N.caninum, не имеет апиразу активности, и его субстратная специфичность ограничивается нуклеозидтрифосфат (Асаи и др., 1998).