В процесс биосинтеза белка рибосомами, называемого трансляцией, вовлечено множество макромолекул и макромолекулярных комплексов. На этом этапе реализации генетической информации происходит считывание генетической информации, заключенной в мРНК, рибосомами и ее передача полипептидным цепям белков, т.е. биосинтез полипептидных цепей, последовательность аминокислот в которых, как правило, однозначно определена последовательностью нуклеотидов в транслируемых мРНК в соответствии с генетическим кодом. Свободные аминокислоты не узнаются рибосомами. Чтобы это произошло, аминокислоты должны поступать в рибосомы в виде конъюгатов с тРНК (аминоацилированных тРНК), последовательности нуклеотидов которых распознаются аппаратом трансляции. В каждой молекуле тРНК имеется участок из трех нуклеотидов, комплементарный кодону мРНК. Именно эта последовательность, называемая антикодоном, в основном определяет положение той или иной аминокислоты в полипептидной цепи. В ходе каждого индивидуального акта трансляции рибосома распознает кодон мРНК и в соответствии с ним выбирает аминоацилированную тРНК, антикодон которой соответствует транслируемому кодону. После этого происходит соединение посредством пептидной связи очередной аминокислоты с С-концевой аминокислотой растущей цепи полипептида.
Таким образом, во время трансляции рибосома после связывания мРНК начинает последовательно, кодон за кодоном, перемещаться вдоль матрицы, выбирая из окружающей среды молекулы аминоацилированных тРНК. При этом каждый индивидуальный акт трансляции завершается присоединением выбранной молекулы аминокислоты к С-концевой аминокислоте синтезируемой цепи белка посредством пептидной связи. Ниже более подробно будут рассмотрены основные этапы биосинтеза белка и компоненты белоксинтезирующей системы бактерий.
Рибосомы
Рибосомы представляют собой крупный рибонуклеопротеидный комплекс с молекулярной массой ~ 2,5 мДа, состоящий из рибосомных белков, молекул рРНК и ассоциированных с ними факторов трансляции. Рибосомы прокариотических и эукариотических организмов различаются по размерам. У эукариот они представлены 80S частицами, тогда как коэффициент седиментации рибосом прокариот составляет 70S. Рибосомы всех известных организмов построены из двух неравных субчастиц: прокариотические – 30S и 50S, а эукариотические – 40S и 60S. 70S рибосомы эубактерий в своем составе содержат 55–60 рибосомных белков, для 80S рибосом эукариот это число составляет 75–85. В обоих случаях рибосомные белки в составе рибосом ассоциированы с молекулами рРНК, образуя пространственно организованные рибонуклеопротеиновые тяжи.
Рибосомные белки E. coli. В настоящее время более 50 рибосомных белков выделено в высокоочищенном состоянии. Молекулярная масса самого маленького белка составляет 5 кДа, а самого большого – 61 кДа, тогда как для большинства рибосомных белков эти значения лежат в пределах 10–20 кДа. Определены аминокислотные последовательности полипептидных цепей всех рибосомных белков E. coli. Малая рибосомная субчастица содержит 21 белок с суммарной молекулярной массой 350 кДа.
Белки в составе 30S субчастицы ассоциированы с 16S РНК, длина которой составляет 1542 нуклеотида (нт). Суммарные молекулярные массы малой и большой субчастиц рибосом достигают соответственно 850 и ~ 1450 кДа. Третья часть массы большой субчастицы приходится на 34 рибосомных белка, а две третьих – на 23S (2904 нт) и 5S рРНК (120 нт). Продолжают накапливаться биохимические данные, указывающие на центральную, возможно ключевую, роль рРНК в обеспечении этапов трансляции. Обнаружены специфические внутримолекулярные и межмолекулярные взаимодействия между различными функциональными участками рРНК. На прямое участие 23S рРНК в трансляции указывает наличие специфических комплементарных взаимодействий между ней и CCA-концами тРНК, акцептирующими аминокислотные остатки. В уточненных пространственных моделях 30S и 50S субчастиц, рассмотренных ниже, сегменты рРНК со специфическими структурными особенностями располагаются в функционально значимых участках рибосом.
Методы исследования пространственной структуры рибосом. Вся информация о пространственной структуре рибосом получена с использованием современных методов, в которых реализуются два направления исследований: структурно-биологические исследования низкого и высокого разрешения, а также биохимические – высокого разрешения. Получаемые результаты, по мере совершенствования методов, все более сближаются друг с другом.
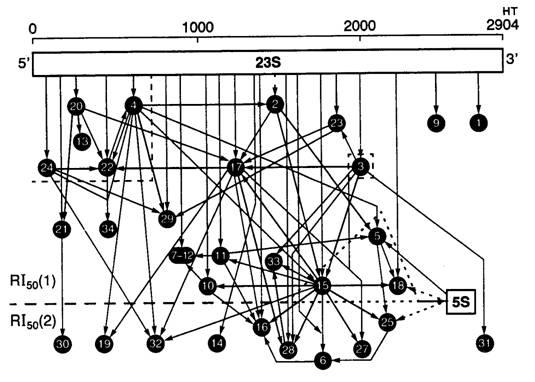
Рис. I.17. Карта сборки 50S субчастицы рибосом E. coli из индивидуальных рибосомных белков и рРНК в процессе реконструкции in vitro
Стрелками обозначены межмолекулярные взаимодействия, удерживающие индивидуальные белки (цифры в кружках) в составе субчастицы, и взаимозависимость сборки. Толщина стрелок отражает прочность связей
Одними из первых следует упомянуть кристаллографию и ЯМР-спектроскопию. Имеются значительные достижения в изучении пространственной структуры небольших субдоменов рРНК с помощью ЯМР-спектроскопии, а также в исследовании структуры индивидуальных рибосомных белков с использованием методов ЯМР и кристаллографии. В последнее время закристаллизованы отдельные рибосомные субчастицы и 70S рибосомы галофильных и термофильных бактерий. Получены картины дифракции для 50S субчастиц галофилов с разрешением 3 Å, а также рибосом и субчастиц T. thermophilusс разрешением 7–20 Å. Однако эти картины дифракции, по крайней мере, в 10 раз сложнее тех, с которыми приходилось иметь дело раньше при расшифровке структур несимметричных макромолекул, что сильно затрудняет их интерпретацию. В результате наиболее распространена в настоящее время альтернативная стратегия определения структуры рибосом "по частям": через пространственные структуры индивидуальных рибосомных белков, субдоменов рРНК и их небольших комплексов. Выявленное в ходе этих исследований большое разнообразие пространственных структур рибосомных белков указывает на существование не менее разнообразных механизмов распознавания белками специфических участков РНК, количество которых должно значительно превышать ограниченное число известных механизмов ДНК–белкового узнавания.
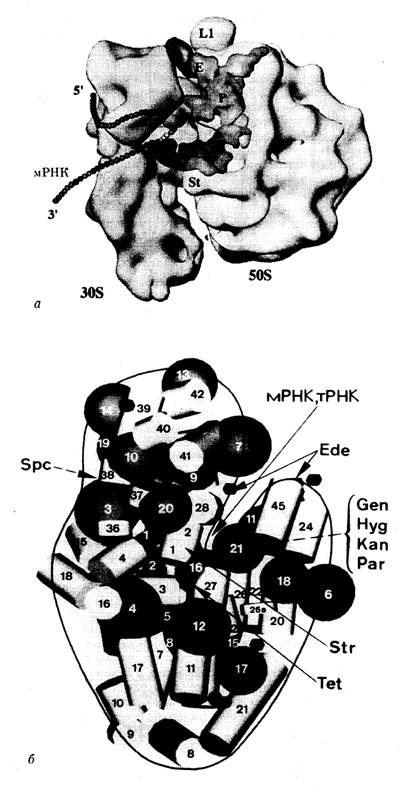
Рис. I.18. Полупрозрачная модель пространственной структуры рибосомы E. coli (а) и расположение рибосомных белков, а также рРНК в ее 30S-субчастице (б)
Указано положение мРНК, тРНК в А-, Р- и Е-участках рибосомы, белка L1 и "выступа" (stalk) 50S субчастицы. Черными шарами обозначены рибосомные белки, светлыми цилиндрами – спиральные участки 16S рРНК, стрелками – места связывания мРНК, тРНК и антибиотиков, цифрами – рибосомные белки и участки рРНК согласно общепринятой номенклатуре
Другим плодотворным направлением исследования пространственной структуры рибосом при низком разрешении является электронная микроскопия (ЭМ). При этом образцы с рибосомами быстро замораживают в жидком этане в тонком слое буфера и исследуют с помощью ЭМ при температуре жидкого азота и низких дозах облучения для сохранения чувствительных к радиации структур в интактном состоянии. На получаемых в результате микрофотографиях могут одновременно содержаться сотни и тысячи по-разному ориентированных индивидуальных рибосом, изображения которых далее подвергаются компьютерному анализу с последующей реконструкцией трехмерной структуры индивидуальной рибосомной частицы. Реконструкцию трехмерных структур работающих рибосом получают в результате анализа микрофотографий ультратонких срезов отдельных бактериальных клеток, активно синтезировавших белок или находившихся в состоянии блока трансляции. Одной из разновидностей ЭМ, активно использующейся для исследования пространственной структуры рибосом, является иммуноэлектронная микроскопия. Первичные и пространственные структуры рибосомных белков, формирующие их эпитопы, значительно различаются, поэтому такие белки редко дают перекрестные иммунологические реакции и их можно четко идентифицировать с помощью специфических антител. При анализе комплексов антител с рибосомами с помощью электронной микроскопии можно видеть, что многие белки локализованы на поверхности рибосомных субчастиц. Оказалось, что пространственное расположение большинства рибосомных белков весьма консервативно. В частности, у грамположительных и грамотрицательных бактерий гомологичные белки занимают одни и те же места на поверхности рибосомных субчастиц.
Важная информация о расположении индивидуальных рибосомных белков в составе рибосом получена и с помощью метода поперечных сшивок. Используя бифункциональные реагенты, например диэпоксибутан или 2-иминотиолан, осуществляют ковалентное соединение рибосомных белков, расположенных по соседству на расстоянии 5–10 Å друг от друга. Поскольку в настоящее время аминокислотные последовательности всех рибосомных белков известны, этим методом можно однозначно определять, какие аминокислотные остатки в соседних белках участвуют в образовании поперечных сшивок.
Целостность рибосом в водных растворах в значительной степени зависит от температуры и ионных условий, особенно от концентрации двухвалентных ионов (Mg2+ и Ca2+). Понижение концентрации ионов Mg2+ приводит вначале к диссоциации рибосом на большую и малую субчастицы, а затем к последовательному упорядоченному освобождению рибосомных белков из субчастиц вплоть до их полного распада с образованием пула отдельных белков и рРНК. Процесс разборки рибосом обратим, и при восстановлении ионных и температурных условий в реакционной смеси возможно реконструирование рибосомных субчастиц из отдельных компонентов с образованием полноценных функционально активных рибосом. На рис. I.17 представлена карта сборки большой субчастицы рибосом E. coli из отдельных компонентов, которая отражает последовательность присоединения рибосомных белков к рРНК, а также два основных этапа сборки. Для перехода ко второму этапу необходимо дальнейшее изменение ионных условий и температуры реакционной среды. Процесс сборки субчастиц рибосом является кооперативным, т.е. присоединение одних рибосомных белков стимулирует включение других. При этом белки, включающиеся в состав рибосомных субчастиц друг за другом, в зрелых субчастицах оказываются расположенными рядом. Реконструирование рибосом из отдельных компонентов in vitro внесло большой вклад в понимание пространственной организации рибосомных субчастиц и функциональной значимости отдельных рибосомных белков.
Метод рассеяния нейтронов на протонах, входящих в состав белков, также способствовал пониманию пространственного расположения белков в рибосомах. Отклонения нейтронов после контактов с протонами белков можно легко отличить от отклонений, которые являются результатом взаимодействия нейтронов с другими атомами, в частности тяжелыми изотопами водорода (2H или 3Н). Если в состав дейтерированных рибосомных субчастиц ввести два рибосомных белка, содержащих обычные протоны, то по рассеянию нейтронов на протонах, характер которого значительно отличается от такового на дейтронах, можно определить расстояния между центрами масс этих двух белков. Усовершенствованный метод нейтронного рассеяния позволяет определять не только расстояния между рибосомными белками, но и пространственную организацию самих полипептидных цепей в составе рибосомных субчастиц. Полученные таким образом нейтронные карты основаны на измерении расстояний между 93 белками. Такие карты имеют фундаментальное значение в интерпретации экспериментальных данных, полученных другими методами, особенно в результате молекулярного моделирования.
При отсутствии данных рентгеноструктурного анализа высокого разрешения молекулярные биологи традиционно обращаются к молекулярному моделированию пространственных структур. В некоторых случаях такой подход бывает весьма успешным, что особенно ярко проявилось при расшифровке пространственной структуры ДНК. Все модели, описанные в настоящее время, учитывают филогенетические особенности вторичной структуры 16S рРНК, и в некоторых из них принимаются во внимание третичные взаимодействия внутри этих макромолекул. В последнее время для таких целей все чаще используется компьютерный анализ. Применение вычислительной техники сводит к минимуму субъективизм в построении моделей и позволяет систематически исследовать возможные конформационные состояния анализируемых молекулярных объектов. При этом выбор конкретного алгоритма в современном моделировании оказывает меньшее влияние на конечный результат, чем выбор имеющихся экспериментальных данных и использование ограничивающих условий.
На рис. I.18 представлена современная модель пространственной структуры 70S рибосомы E. coli, разработанная в лаборатории Д. Франка (США) с учетом данных, которые были получены с помощью всех вышеперечисленных методов.
Этапы биосинтеза белка
Хотя построение первых моделей механизмов биосинтеза белка было начато еще в начале 1960-х гг., полное описание процесса трансляции далеко до завершения и в настоящее время. Ниже будут кратко рассмотрены основные черты классической модели биосинтеза белка рибосомами E. coli, а также особенности некоторых альтернативных моделей.
Процесс биосинтеза белка рибосомами, как и биосинтез любой другой макромолекулы клетки, условно разделяют на три основных этапа: инициацию, элонгацию и терминацию. Во время инициации трансляции происходит сборка нативной 70S или 80S рибосомы на транслируемой мРНК и подготовка к образованию пептидной связи между первыми двумя N-концевыми аминокислотными остатками синтезируемого полипептида. При элонгации наблюдается последовательное удлинение растущей цепи полипептида аминокислотными остатками, а терминация трансляции сопровождается прекращением синтеза полипептида и его высвобождением из трансляционного комплекса. При этом наблюдается также разделение рибосомы и мРНК, после чего они вступают в новый цикл трансляции. В ходе трансляции рибосома последовательно перемещается вдоль транслируемой молекулы мРНК, считывая заключенную в ней генетическую информацию в виде триплетного генетического кода. Трансляция начинается в 5’-концевой части мРНК, а завершается в ее 3’-концевой части. При этом биосинтез полипептида начинается с его N-концевой аминокислоты. Рассмотрим каждый из вышеперечисленных этапов более подробно на примере белоксинтезирующей системы E. coli.
Инициация трансляции. Биосинтез белка рибосомами начинается с образования комплекса между малой 30S субчастицей рибосом, инициаторной тРНК и участком транслируемой мРНК, содержащим сайт связывания рибосом, который включает в себя инициирующий (как правило, AUG) кодон. В образовании инициационного комплекса с 30S субчастицей принимают участие три белковых фактора инициации – IF1, IF2 и IF3. В ходе этого процесса расходуется одна молекула GTP, которая взаимодействует с IF2 и изменяет его конформацию (см. рис. I.19). Таким образом, на первом этапе образования инициационного комплекса происходит объединение свободной 30S субчастицы с факторами инициации и GTP, после чего с ними последовательно связываются мРНК и инициаторная тРНК (в случае E. coli, как правило, формилметионил(fMet)-тРНКfMet). Инициаторная тРНК строго специфична для этой стадии белкового синтеза. Сначала она обычным путем акцептирует Met с образованием Met-тРНКfMet, а затем специальная ферментная система E. coli формилирует NH2-группу остатка Met. Последовательность присоединения инициаторной тРНК и мРНК к 30S субчастице не имеет значения, что и отражено на рис. I.19 (стадии А, А’, В, В’).
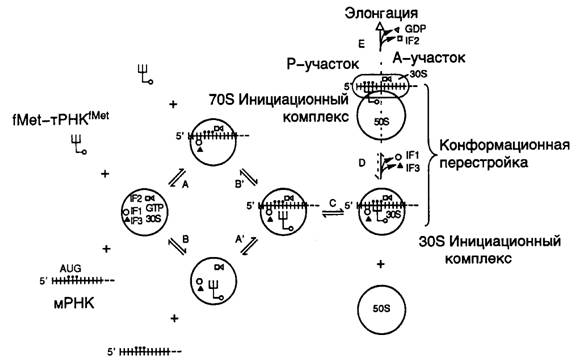
Рис. I.19. Этапы инициации трансляции у E. coli
Вначале после объединения факторов инициации трансляции, GTP, fMet-тРНКfMet и мРНК с 30S субчастицей антикодон тРНК еще не взаимодействует с инициаторным AUG-кодоном (стадии А’ и B’). Такое продуктивное взаимодействие тРНК с мРНК происходит в дальнейшем (стадия C), и этот переход является одной из лимитирующих стадий всего процесса образования инициационного комплекса. С завершением стадии C происходит формирование стабильного тройного (из трех основных компонентов) инициационного комплекса, сопровождаемое конформационными перестройками всех его компонентов. После выхода из комплекса факторов инициации трансляции IF1 и IF3 тройной комплекс приобретает способность связывать большую 50S субчастицу рибосом, что сопровождается дальнейшими конформационными перестройками всей рибосомы (стадия D). В ходе этого процесса происходит расщепление молекулы GTP до GDP и ортофосфата и освобождение из комплекса фактора IF2 (стадия E). Формилметионил-тРНКfMet вместе с инициирующим AUG-кодоном перемещаются в донорный (P) участок рибосомы, освобождая акцепторный (A) участок для следующей аминоацилированной тРНК. В результате инициационный комплекс становится полностью подготовленным для вступления в следующую фазу биосинтеза белка – элонгацию полипептидных цепей.
Элонгация. В соответствии с обсуждаемой моделью принято считать, что после образования тройного комплекса, включающего 70S рибосому, мРНК и инициаторную тРНК, завершается этап инициации трансляции, и процесс биосинтеза белка вступает в фазу элонгации, которая завершается освобождением полипептидных цепей из элонгирующих комплексов. Во время элонгации происходит последовательное присоединение аминокислотных остатков к C-концевым частям строящихся полипептидных цепей, направляемое кодонами транслируемых матричных РНК.
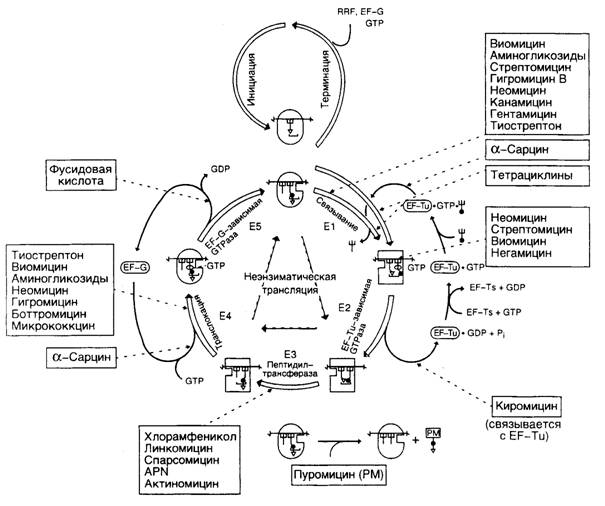
Рис. I.20. Элонгация полипептидных цепей
Стрелки указывают на реакции, подавляемые соответствующими антибиотиками
Этап элонгации начинается со взаимодействия фактора элонгации трансляции EF-Tu, молекулы GTP и очередной аминоацилированной тРНК с A-участком рибосомы (см. рис. I.20, стадия Е1). Вхождение аминоацилированной тРНК в A-участок происходит в соответствии с установленным в нем кодоном транслируемой мРНК. При этом лишь та аминоацилированная тРНК прочно связывается с рибосомой, у которой антикодон комплементарен кодону, установленному в A-участке. После гидролиза GTP и освобождения EF-Tu•GDP из комплекса (стадия Е2) происходит образование новой пептидной связи между карбоксильной группой формилметионина инициаторной тРНК и NH2-группой аминокислотного остатка, находящегося в A-участке рибосомы в составе аминоацил-тРНК (стадия Е3). Эта стадия получила название транспептидации. Обмен GDP на GTP в освободившемся комплексе EF-Tu•GDP происходит с участием фактора EF-Ts.
Образовавшийся в итоге пептид удерживается рибосомой через остаток тРНК, находящийся в A-участке, а освободившаяся тРНК временно сохраняется в так называемом E-участке рибосомы (от англ. exit – выход). Такая соединенная с пептидом тРНК получила название пептидил-тРНК. Образовавшаяся пептидил-тРНК далее переносится из A- в P-участок рибосомы. Эта стадия элонгации (Е4) известна под названием транслокации. Транслокация индуцируется фактором элонгации EF-G, который освобождается из элонгирующего комплекса после расщепления молекулы GTP. Таким образом, энергия еще одной молекулы GTP используется в акте транслокации. После завершения транслокации происходит освобождение фактора EF-G из элонгирующего комплекса. При этом A-участок рибосомы остается свободным. Следующий цикл элонгации начинается с вхождения в A-участок рибосомы в составе тройного комплекса очередной молекулы тРНК (стадия Е1), что сопровождается освобождением формилметионил-тРНКfMet из E-участка, после чего повторяются все остальные вышеперечисленные стадии элонгации. В физиологических условиях рибосома совершает ~ 20 циклов элонгации в секунду. В соответствии с этим для синтеза белка длиной в 200 аминокислотных остатков требуется ~10 секунд.
В рассмотренной классической модели биосинтеза белка с тремя участками связывания тРНК на любой стадии элонгации с рибосомой взаимодействуют две молекулы тРНК. Иными словами, до стадии транслокации тРНК занимают A- и P-участки рибосомы, тогда как после транслокации молекулы ассоциированы с P- и E-участками. Между участками A и E существует аллостерическое взаимодействие, что проявляется в отрицательном кооперативном эффекте связывания молекул тРНК этими участками и означает, что только A- или E-участки рибосомы могут быть заняты молекулой тРНК, и рибосома не содержит одновременно занятыми оба участка.
Молекулярная мимикрия фактора элонгации EF-G. Недавно было обнаружено, что пространственная структура домена IV полипептидной цепи фактора элонгации EF-G имитирует структуру тРНК в ее комплексе с другим фактором элонгации EF-Tu. При этом структура соответствующей части полипептидной цепи EF-G напоминает форму антикодоновой петли тРНК в комплексе с фактором элонгации, ее положение относительно к ó ровой части EF-Tu и даже распределение электростатических зарядов на поверхности полипептида, которое соответствовало таковому углевод-фосфатного остова тРНК. Это открытие позволило по-новому посмотреть на механизм действия EF-G в цикле трансляции. Такого рода данные позволили предположить, что домен IV фактора EF-G занимает во время некоторых этапов транслокации ту же часть A-участка рибосом, что и тРНК. Однако остается непонятным, каким образом это может физически влиять на прохождение акта транслокации.
Терминация трансляции. Процесс трансляции вступает в завершающую фазу после того, как в A-участок рибосомы попадает терминирующий (бессмысленный) кодон мРНК, а пептидил-тРНК перемещается в донорный P-участок рибосомы. Белковые факторы RF1 и RF2 участвуют в распознавании последовательностей нуклеотидов терминирующего кодона. Фактор RF3, также как и EF-G, имитирует структурные особенности фактора EF-Tu, что дает ему возможность взаимодействовать с А-участком рибосомы. Но поскольку с ним не связана аминоацилированная тРНК, его присутствие в А-участке приводит к обрыву строящейся цепи полипептида. После формирования такого комплекса происходит расщепление сложноэфирной связи между полипептидом и тРНК, а также освобождение синтезированного полипептида. С помощью мутационного анализа было установлено, что молекулы рРНК обеих субчастиц рибосом E. coliучаствуют в гидролизе пептидил-тРНК.
Для того чтобы рибосома оставшегося комплекса рибосома–мРНК–тРНК могла вступить в следующий цикл трансляции, она должна освободиться из него. Установлено, что вышеупомянутые рибосомные рилизинг-факторы (RF) совместно с фактором EF-G при участии молекулы GTP обеспечивают диссоциацию комплекса на составные компоненты, которые затем вступают в новый раунд белкового синтеза. Фактор RF4 (иначе называемый RRF – ribosome-recycling factor) не имеет аналога у эукариот. Его роль, по-видимому, заключается в стимуляции перемещения молекулы деацилированной тРНК из P-участка в E-участок рибосомы и(или) удалении оставшегося RF-фактора из A-участка. Это способствует полному освобождению рибосомы и ее вовлечению в новый цикл трансляции в результате инициации или реинициации синтеза белка. Отделившаяся от мРНК рибосома перед вступлением в новый цикл диссоциирует на две субчастицы под действием фактора инициации трансляции IF3. Альтернативно, в том случае, если новый инициирующий кодон полицистронной матрицы находится достаточно близко от стоп-кодона, синтез белка может быть реинициирован.
Реинициация трансляции. Реинициацией трансляции называют повторное вступление рибосом, терминировавших биосинтез белка, в цикл трансляции без предварительного отделения их от мРНК. Реинициация синтеза белка широко распространена у E. coliи играет важную роль в контроле экспрессии генов этого микроорганизма на уровне трансляции. Это связано с тем, что значительная часть бактериальных мРНК полицистронна, а, следовательно, за терминирующим кодоном одного цистрона на небольшом от него удалении располагается инициирующий кодон следующего. Благодаря реинициации имеет место координированная (сопряженная) трансляция нескольких ОРС, объединенных в полицистронной матрице.
Имеются данные о том, что рибосомы E. coli, терминировавшие синтез полипептида, обладают способностью перемещаться на короткие расстояния в окрестностях терминирующего кодона и после встречи с инициирующим кодоном в новом сайте инициации трансляции могут начать следующий раунд трансляции без отделения от мРНК. Новый инициирующий кодон может располагаться выше или ниже стоп-кодона предыдущего гена, а может и перекрываться с ним (например, в последовательности AUGA). Реинициация может происходить с полной эффективностью, если в сайте реинициации имеется адекватная SD-последовательность, а терминирующий и инициирующий кодоны расположены достаточно близко друг к другу (менее эквивалента длины рибосомы). Последнее обстоятельство указывает на быструю кинетику отделения терминировавших рибосом от мРНК.
Важным следствием сопряжения трансляции у прокариот через реинициацию является зависимость экспрессии целой серии генов, принадлежащих одному оперону, от трансляции первой ОРС полицистронной матрицы. При этом следует заметить, что сопряжение экспрессии генов на уровне трансляции может оказывать и негативное влияние на эффективность трансляции нижерасположенных цистронов.
Если обычная инициация трансляции у E. coli на кодоне UUG происходит с очень низкой эффективностью, то реинициирующие рибосомы используют его для начала синтеза белка весьма охотно. Это объясняют отсутствием фактора IF3 в реинициирующем комплексе, который во многом определяет точность выбора инициирующего кодона рибосомами.
На поведение рибосом, терминировавших синтез белка, большое влияние оказывают и факторы терминации трансляции (RF). В опытах с мутантным бактериофагом R17, содержащим амбер-кодон в положении 7 гена белка оболочки, было установлено, что в отсутствие фактора терминации трансляции RF4 вслед за терминацией трансляции на бессмысленном кодоне имела место реинициация трансляции на следующем кодоне мРНК, что завершалось синтезом белка оболочки, укороченного с N-конца на 7 аминокислот. На этом основании полагают, что рилизинг-фактор RF4 в обычных условиях предотвращает распознавание аминоацилированной тРНК кодона, находящегося в А-участке рибосомы, который в мРНК следует за терминирующим.
Альтернативные модели цикла трансляции. В рассмотренной выше классической модели трансляции перемещение молекул тРНК на большой и малой субчастицах рибосом сопряжено друг с другом. В отличие от этого, в активно обсуждающейся модели гибридных состояний (hybrid states model) перемещение тРНК между A- и P-участками 30S субчастицы происходит независимо от перемещения тРНК между A-, P- и E-участками большой субчастицы. В соответствии с этой моделью аминоацил-тРНК попадает в пептидил-тРНК–рибосомный комплекс в составе тройного комплекса EF-Tu·GTP·тРНК и взаимодействует с ней первоначально в гибридном состоянии A/E. В этом состоянии антикодоновая часть тРНК связывается с A-участком 30S субчастицы, а ее CCA-конец, удерживаемый EF-Tu, располагается в E-участке большой субчастицы и, возможно, частично на малой субчастице. Вслед за гидролизом GTP происходит освобождение EF-Tu, что делает возможным перемещение CCA-конца аминоацил-тРНК в A-участок большой субчастицы, приводящее к возникновению A/A-состояния, эквивалентного состоянию взаимодействия аминоацил-тРНК с A-участком в классической модели. После образования пептидной связи аминоацил-тРНК, уже связанная с растущей полипептидной цепью, перемещается в P-участок большой субчастицы, а деацилированная тРНК переходит в E-участок большой субчастицы. Вновь образованная пептидил-тРНК находится теперь в гибридном A/P-состоянии: антикодоновая часть остается в A-участке 30S субчастицы, а CCA-конец занимает P-участок большой субчастицы рибосом. При этом деацилированная тРНК находится в гибридном P/E-состоянии: антикодоновый конец остается в P-участке малой субчастицы, тогда как CCA-конец занимает E-участок большой субчастицы. Далее фактор элонгации EF-G в GTP-зависимой реакции обеспечивает перемещение антикодоновой части тРНК, находящейся в гибридном состоянии, вместе с мРНК относительно 30S субчастицы. При этом пептидил-тРНК переходит в чувствительное к пуромицину P/P-состояние, соответствующее ее взаимодействию с P-участком в классической модели, а деацилированная тРНК находится в E-состоянии и на этом этапе трансляции может взаимодействовать только с E-участком большой субчастицы рибосом.
По крайней мере, три интересных следствия вытекают из модели гибридного состояния. Во-первых, пептидильная часть растущего пептида остается на рибосомах в стационарном состоянии, а во время трансляции перемещается тРНК. Во-вторых, транслокация тРНК происходит в два этапа: во время первой стадии обе молекулы тРНК движутся относительно большой субчастицы, на втором этапе обе молекулы тРНК вместе со связанной с ними мРНК перемещаются относительно малой 30S субчастицы рибосом. В-третьих, в процессе синтеза белка имеют место не два или три состояния связывания тРНК, а шесть или даже, возможно, семь таких состояний.
С помощью физических методов были получены прямые доказательства спонтанного прохождения стадии транслокации, опосредуемой пептидилтрансферазой рибосом. При использовании флуоресцентных зондов, связанных с различными участками тРНК и рибосом, удалось обнаружить изменения в квантовом выходе флуоресценции и анизотропные эффекты при образовании пептидной связи, что указывало на перемещение молекулы тРНК относительно рибосомных белков S21 и L11. На основании этих данных было высказано предположение, что во время пептидилтрансферазной реакции пептидильная цепь остается в постоянном положении относительно рибосомы, а перемещаются молекулы тРНК. Эта модель пептидилтрансферазной реакции получила название модели перемещения (displacement model). Она обладает многими общими чертами с моделью гибридного состояния, однако отличается тем, что в этой модели движение мРНК в пептидилтрансферазной реакции сопровождает перемещение тРНК.
Замечания о точности трансляции. Сама по себе стабильность кодон–антикодоновых взаимодействий не может обеспечивать наблюдаемую высокую точность трансляции. Рибосомы активно участвуют в акте распознавания молекулами тРНК соответствующих кодонов мРНК, повышая точность функционирования этого механизма, по крайней мере, на четыре порядка. Данный эффект объясняют функционированием механизмов, корректирующих ошибки на этом этапе трансляции, которые сопряжены с EF-Tu-зависимым гидролизом GTP во время выбора соответствующей аминоацилированной тРНК. Недавние измерения скорости гидролиза GTP рибосомами в присутствии правильной (cognate) или неправильной (noncognate) тРНК и искусственной матрицы показали, что в первом случае она выше в ~ 104 раз. Это приводит к преимущественному освобождению комплекса EF-Tu·GDP из рибосом, содержащих правильные аминоацил-тРНК в А-участке. Для реализации данного механизма рибосомы должны распознавать правильные и ошибочные кодон-антикодоновые взаимодействия, а также передавать эту информацию своему GTPазному центру.
Мутантные рибосомы, для которых характерна пониженная точность трансляции, как правило, обладают более высоким сродством к тРНК в A-участке. Напротив, у "сверхточных" рибосом такое сродство понижено. В соответствии с этим повышенную точность трансляции можно объяснять в терминах уменьшения неспецифического связывания аминоацилированных тРНК A-участком рибосом и vice versa. Недавно было показано, что сродство тРНК к P-участку таких мутантных рибосом изменяется на противоположное таковому A-участка: у ram-мутантов с низкой точностью трансляции наблюдается пониженное сродство P-участка к тРНК, а у "сверхточных" рибосом это сродство повышено. Таким образом, в настоящее время полагают, что простые реципрокные отношения связывают A- и P-участки рибосом с механизмами, которые управляют взаимодействием мРНК и соответствующих тРНК с рибосомами.
Новая 10Sa РНК, функционирующая в трансляции. В том случае, если транслируемая бактериальная мРНК укорочена в своей 3’-концевой части и рибосома не находит в соответствующей рамке считывания терминирующий кодон, она не может закончить трансляцию с помощью стандартного механизма терминации. Полагают, что, столкнувшись с такой ситуацией, рибосома останавливается на конце мРНК и с ней взаимодействует недавно открытая 10Sa РНК, кодируемая у E. coli геном ssrA, которая запускает процесс деградации частично синтезированного полипептида. Эта необычная РНК, аминоацилированная остатком Ala на своем 3’-конце, подобном стандартной акцепторной последовательности тРНК, сочетает в себе свойства тРНК и мРНК. 10Sa РНК, как и тРНКAla, образует внутреннюю пару оснований G3:U70, которая необходима для специфического распознавания тРНКAla рибосомами. После взаимодействия с рибосомой эта РНК вступает в стандартный цикл трансляции: ее остаток Ala включается в недостроенную цепь полипептида. Вслед за этим рибосома транслирует короткую последовательность нуклеотидов 10Sa РНК, которая теперь функционирует в качестве матрицы, добавляя десять аминокислот – ANDENYALAA – в C-конец укороченной полипептидной цепи, и терминирует трансляцию на UAA-кодоне 10Sa РНК. C-Концевая олигопептидная последовательность освободившегося полипептида далее распознается специфической протеиназой, которая расщепляет этот полипептид, обеспечивая его утилизацию бактериальной клеткой в процессе катаболизма.