Векторы, пригодные для клонирования ДНК в бактериях, отличающихся от E. coli, должны обладать всеми характерными чертами, которые были отмечены выше. От только что рассмотренных они отличаются главным образом тем, что содержат природные или искусственные генетические элементы, функционирующие в новых клетках-хозяевах.
Интегрирующие векторы грамположительной бактерииBacillus subtilis. B. subtilis, как и E. coli, является излюбленным объектом генной инженерии. Это связано с тем, что B. subtilis – непатогенный микроорганизм, многие штаммы которого широко используются в микробиологической промышленности для производства биологически активных соединений и пищевых веществ. В отличие от E. coli B. subtilis способна секретировать белки и пептиды, что облегчает их очистку и дальнейшее использование. Большинство векторов для клонирования ДНК в клетках B. subtilis создано на основе плазмид других видов бацилл, а также Staphylococcus aureus и бактерий рода Streptococcus.
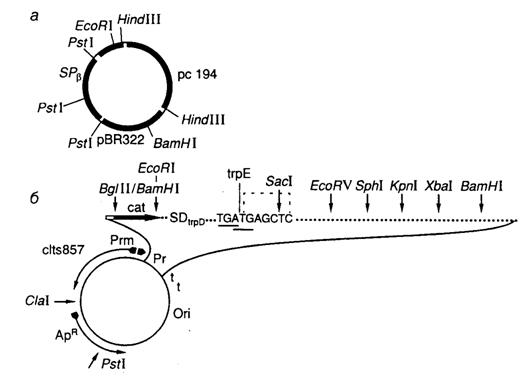
Рис. II.11. Челночный вектор pFH7 (а) и экспрессирующий вектор pPR-TGATG-1 (б)
Обозначено расположение генов и уникальных сайтов рестрикции
Основным свойством интегрирующих векторов является их способность стабильно встраиваться в геном клетки-хозяина. Это становится возможным благодаря наличию в таких векторах последовательностей нуклеотидов, гомологичных последовательностям геномной ДНК. В результате функционирования общей системы рекомбинации происходит объединение хромосомной и плазмидной ДНК интегрирующего вектора, которое приводит к стабильному включению всей векторной плазмиды в хромосому. Примером такого интегрирующего вектора служит плазмида pFH7 B. subtilis (рис. II.11, а). Векторная плазмида содержит фрагмент ДНК умеренного бактериофага SPb и после попадания в клетки B. subtilis, лизогенные по данному бактериофагу, эффективно интегрируется в профаг. Поскольку плазмида содержит ген устойчивости к хлорамфениколу cat, клетки приобретают этот признак. Индукция профага приводит к образованию фаговых частиц, трансдуцирующих такую плазмиду и ассоциированный с ней признак устойчивости к хлорамфениколу. Интеграция плазмиды SPb в бактериальную хромосому происходит по механизму гомологичной рекомбинации с участием гена recE.
Способность к интеграции в бактериальную хромосому обнаруживают и другие плазмиды, содержащие фрагменты хромосомной ДНК клеток-хозяев, что продемонстрировано, в частности, для плазмид E. coli и Streptococcus pneumoniae.
Интегрирующие векторные системы, в которых используется тот же принцип гомологичной рекомбинации, разработаны и для эукариотических клеток, включая клетки животных и растений. В конце концов, такие работы привели к развитию целого направления исследований по созданию трансгенных животных и растений, стабильно наследующих и экспрессирующих гены, искусственно введенные в их геном. О некоторых важных следствиях этого направления исследований, включая генотерапию, речь пойдет в главе 10.
Наличие феномена гомологичной рекомбинации между хромосомной ДНК клетки-хозяина и векторной ДНК, содержащей гомологичные хромосомной ДНК последовательности нуклеотидов, приходится учитывать при получении соответствующих генно-инженерных конструкций. Такая неконтролируемая рекомбинация в большинстве случаев нежелательна, так как может приводить к потере или структурным перестройкам клонированных фрагментов ДНК. Для того чтобы свести последствия этого явления к минимуму, используют специальные штаммы клеток-хозяев, в которых общая система рекомбинации не функционирует, например вследствие мутационной инактивации гена recA E. coli или recE B. subtilis.
Челночные векторы. Интегрирующая плазмида pFH7 (см. рис. II.11, а) дает возможность проиллюстрировать еще один важный принцип, широко используемый при конструировании векторных систем в генной инженерии. Эта плазмида получена путем объединения двух репликонов, один из которых берет начало от плазмиды pC194 B. subtilis, а другой – от плазмиды pBR322 E. coli, что дает возможность вектору существовать и стабильно реплицироваться как в клетках E. coli, так и B. subtilis. Такие векторы, способные реплицироваться в клетках-хозяевах разных биологических видов, называют челночными, или бинарными векторами.
Принципы конструирования и функционирования челночных векторов одинаковы, они должны включать в себя репликоны тех генетических систем, в которых будет происходить репликация челночного вектора. При этом используются области начала репликации генетических элементов, которые автономно существуют во внехромосомном состоянии в природных условиях. Так, обсуждавшийся выше интегрирующий вектор pFH7 B. subtilis обладает свойствами челночного вектора, поскольку для его конструирования использованы репликоны двух видов бактерий. Более эффектными примерами челночных векторов являются плазмидные ДНК, способные реплицироваться в клетках высших (животных и растений) и низших организмов. Необходимость использования челночных векторов в генной инженерии связана с тем, что наработку в препаративном количестве векторной ДНК для проведения генно-инженерных манипуляций удобнее проводить в бактериальных клетках, тогда как получение биологически активных продуктов клонированных генов высших организмов во многих случаях возможно только в клетках своего или близкого вида, в которых эти гены экспрессируются в природных условиях, т.е. в своем обычном генетическом окружении (подробнее см. раздел 7.6).