Эта линия клеток и ее многочисленные производные часто используются для синтеза рекомбинантных белков после предварительной эндогенной амплификации соответствующих рекомбинантных генов, введенных в клетки с помощью трансфекции. Спонтанная амплификация генов в нормальных клетках животных является редким событием, а в трансформированных (опухолевых) клетках она происходит с частотой 10-4–10-6 за клеточную генерацию. Частота амплификации определенных генов может быть повышена в результате некоторых экзогенных воздействий, включая инкубацию клеток с гидроксимочевиной, афидиколином, канцерогенами, а также под действием ультрафиолетового света или g-излучения и в условиях гипоксии. Амплификация гена обычно начинается с его вырезания из хромосомы в составе протяженного сегмента ДНК, сопровождаемого образованием кольцевой структуры, которая продолжает существовать в виде внехромосомного элемента (двойной минихромосомы (double-minutes – DM)), способного к автономной репликации. Предполагают, что DM является промежуточным продуктом амплификации, образующимся в условиях низкого давления селектирующего фактора окружающей среды. На более поздних стадиях амплифицирующаяся ДНК после повторной интеграции в геном обнаруживается преимущественно в составе удлиненного участка хромосомы (extended chromosomal region – ECR), иначе называемого гомогенно окрашивающейся областью (homogeneously staining region – HSR). На этой стадии амплифицированный участок генома клеток, а следовательно, и их фенотип становятся более стабильными.
Для биотехнологических целей в системе амплификации рекомбинантных генов в качестве селектируемого маркера часто используют ген дигидрофолатредуктазы (ДГФР), интегрированный в экспрессирующий вектор. ДГФР катализирует превращение фолиевой кислоты в тетрагидрофолат, необходимый для синтеза эукариотическими клетками глицина, тимидинмонофосфата и пуриновых оснований. В этой связи клетки CHO, в которых ген ДГФР инактивирован под действием мутации, не растут на средах без нуклеозидов и приобретают эту способность после трансфекции геном ДГФР. Растущие трансфектанты далее отбираются по признаку амплификации гена ДГФР на фоне увеличивающихся концентраций метотрексата – ингибитора ДГФР, в питательной среде, так как увеличение числа копий гена будет придавать клеткам устойчивость к ингибитору в больших концентрациях. После проведения множественных раундов селекции получают популяцию клеток, содержащих до нескольких сотен копий гена ДГФР.
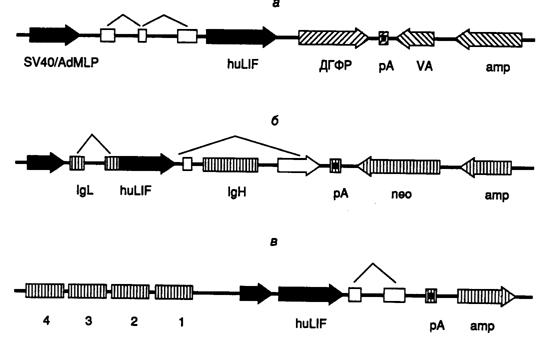
Рис. II.13. Кассеты генов в экспрессирующих векторах, использованные для получения фактора, ингибирующего лейкозы, в культурах клеток
а – кДНК huLIF (черные стрелки), включающая лидерные последовательности (незаштрихованные прямоугольники), была интегрирована в экспрессирующий вектор pXMT3 под контроль гибридного промотора, расположенного между точкой начала репликации вируса SV40 и основным поздним промотором аденовируса (SV40/AdMLP), ДГФР, amp, neo – гены дигидрофолатредуктазы и устойчивости к ампициллину и неомицину соответственно, VA – трансляционный энхансер; pA – сайт полиаденилирования;
б – вектор pSV2neo-Ig-huLIF, сконструированный на основе транскрипционных и трансляционных регуляторных элементов генов тяжелой (IgH) и легкой (IgL) цепей иммуноглобулинов. Транскрипция кассеты контролируется мышиным промотором гена IgL k и энхансером гена IgH. Транскрипт, содержащий последовательность huLIF, в результате сплайсинга объединяется с экзоном 2 гена IgL, кодирующего лидерную последовательность, которая обеспечивает секрецию зрелого huLIF клетками;
в – вектор pGES-LIF, прямоугольники 1–4 – регуляторная область кластера глобиновых генов, короткая черная стрелка – промотор b-глобинового гена, ломаной линией отмечены последовательности, удаляемые сплайсингом. Во всех случаях стрелки указывают направление транскрипции
Размер амплифицированного геномного локуса значительно больше размера селектируемого гена и может составлять несколько сотен тысяч пар оснований. Трансфекция клеток геном ДГФР и исследуемым рекомбинантным геном, заключенными в один экспрессирующий вектор, сопровождаемая их интеграцией в геном, будет приводить к их совместной амплификации на фоне метотрексата и усилению синтеза требуемого рекомбинантного белка, как следствие увеличения дозы его гена. Данные о возможности сохранения высокого уровня экспрессии гена в отсутствие селектирующего агента, т.е. генетической стабильности продуцентов, противоречивы. Результат зависит от генотипа конкретных клонов клеток, в том числе и локализации сайта интеграции экспрессирующего вектора в геном клеток CHO. Эффективным экспрессирующим вектором, используемым в описываемой системе, является двухцистронная молекула ДНК, в которой исследуемый рекомбинантный ген находится под контролем сильного промотора перед геном ДГФР, что обеспечивает высокое сопряжение синтеза рекомбинантного белка с уровнем устойчивости клеток к метотрексату (рис. II.13, а).