Делецией называют потерю части нуклеотидов в геноме организма. Такой вид мутаций удобнее всего использовать для локализации (картирования) функционально значимых участков генов и кодируемых этими генами белков. Действительно, последовательное удаление все новых и новых участков ДНК на границах генов с помощью делеций оказалось исключительно плодотворным в обнаружении регуляторных элементов генов, исследовании их структурно-функциональных особенностей, взаимного расположения и влияния друг на друга. Простой и эффективный метод получения делеций любого размера разработан с использованием экзонуклеазы Bal31 для удаления нуклеотидов в окрестностях сайтов рестрикции (рис. II.16).
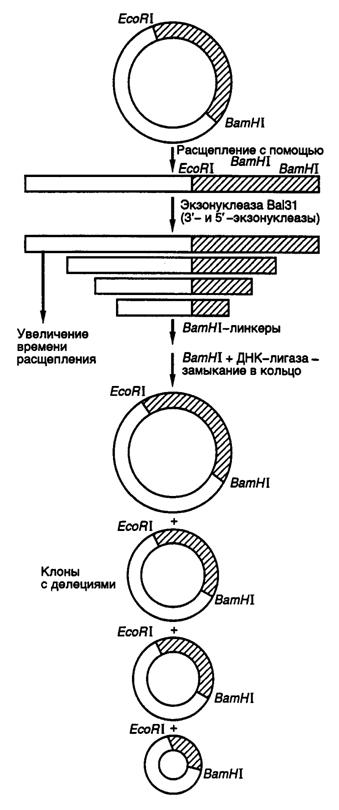
Рис. II.16. Получение делеций с помощью нуклеазы Bal31
Ген, клонированный в плазмиде, расщепляют по уникальному сайту рестрикции и образовавшиеся линейные молекулы ДНК инкубируют в присутствии экзонуклеазы Bal31. При этом экзонуклеаза последовательно удаляет нуклеотиды с обоих концов ДНК, причем количество удаляемых нуклеотидов прямо пропорционально времени инкубации ДНК с нуклеазой, а также зависит от температуры инкубации и концентрации фермента. В результате подобного действия образуется набор фрагментов ДНК разной длины, содержащих делеции различных размеров по обе стороны выбранного сайта рестрикции. К "тупым" концам таких молекул ДНК с помощью ДНК-лигазы присоединяют двухцепочечные олигонуклеотидные линкеры, содержащие уникальный (часто исходный) сайт рестрикции, обрабатывают соответствующей рестриктазой, замыкают молекулы в кольцо посредством лигирования и затем вводят их в бактериальные клетки. Точное картирование концов делеций осуществляют секвенированием соответствующих участков ДНК мутантных плазмид. В результате получают набор делеций разного размера, положение которых в исследуемом фрагменте ДНК строго локализовано.
Небольшие делеции в окрестностях сайтов рестрикции можно получать быстрее удалением "липких" концов линеаризованной плазмидной ДНК после рестрикции с последующим замыканием линейной ДНК в кольцо лигированием по образовавшимся "тупым" концам. В этом случае размер делеции соответствует размеру одноцепочечных "липких" концов в сайтах рестрикции. Кроме того, такой метод допускает простую проверку наличия мутаций в требуемом участке ДНК, так как в результате мутации происходит потеря уникального сайта рестрикции.
Для получения вставок коротких или протяженных последовательностей нуклеотидов в исследуемые участки клонируемых генов также разработаны эффективные и надежные методы. В простейшем случае такая задача решается путем расщепления исследуемого гена рестриктазой по уникальному сайту рестрикции и встраивания по этому сайту фрагмента природной ДНК или синтетического двухцепочечного олигонуклеотида, который фланкирован соответствующими "липкими" концами. Лигирование может проводиться и по "тупым" концам. В таком случае последовательности нуклеотидов на концах вставки уже не имеют существенного значения для встраивания этого фрагмента по сайту рестрикции.
Для создания множественных вставок коротких или протяженных последовательностей нуклеотидов в исследуемых участках ДНК в основном используют два подхода. В первом случае с помощью панкреатической ДНКазы в низких концентрациях в присутствии ионов Mn2+ вносят случайным образом один двухцепочечный разрыв в каждую векторную плазмиду, содержащую клонированный ген. К концам образовавшихся линейных молекул ДНК присоединяют с помощью ДНК-лигазы синтетические олигонуклеотидные линкеры, содержащие сайт рестрикции, который отсутствует в исследуемой плазмиде. Образовавшиеся линейные молекулы ДНК с линкерами обрабатывают рестриктазой, узнающей сайт рестрикции линкера, что приводит к образованию "липких" концов, и замыкают в кольцо с помощью ДНК-лигазы. В итоге кольцевые молекулы ДНК содержат исследуемый клонированный ген, в котором имеется по одной вставке локализованных случайным образом (в соответствии с расположением исходных двухцепочечных разрывов) олигонуклеотидных линкеров. Во втором случае статистические разрывы в двухцепочечной ДНК получают путем частичного (неполного) гидролиза мелкощепящими рестриктазами, которые узнают сайт рестрикции длиной в 4 нуклеотида. Метод получения вставок с использованием синтетических олигонуклеотидных линкеров получил название сканирования линкером.
Химический мутагенез
Делеции и вставки, создаваемые в структурных частях генов, как правило, их инактивируют, особенно в тех случаях, когда такие мутации приводят к сдвигу открытых рамок считывания. Поэтому делеции и вставки in vitro используют главным образом для поиска и изучения регуляторных элементов генов, влияющих на эффективность их экспрессии. Большое значение для исследования функционирования белков имеют методы мутагенеза in vitro, направленные на получение точковых мутаций, следствием которых являются одиночные замены аминокислот в полипептидных цепях.
Распространенным методом введения большого числа точковых мутаций разной локализации в исследуемые части генов in vitro является химический мутагенез одноцепочечных участков рекомбинантных ДНК. Принцип подобных методов заключается в том, что некоторые химические мутагены, такие как бисульфит натрия, гидроксиламин или метоксиламин, действуют только на одноцепочечные участки ДНК. Следовательно, получив молекулы ДНК, содержащие одноцепочечные бреши в исследуемых участках генов, можно с помощью бисульфита натрия дезаминировать остатки цитозина в этих участках, т.е. превратить их в остатки урацила. После достройки цепи такой мутагенизированной молекулы ДНК с помощью фрагмента Кленова ДНК-полимеразы I E. coli происходит замена исходных G‑С-пар на T–U. Затем мутагенизированные молекулы ДНК с помощью трансформации вводят в бактериальные клетки, где по завершении первого раунда репликации в молекуле осуществляется замена остатков U на T и полная замена G–С-пары на A–T, т.е. имеет место обычная транзиция.
Одноцепочечные мутагенизированные участки ДНК удобно получать путем гибридизации одноцепочечной ДНК вектора с двухцепочечной ДНК того же вектора, содержащего клонированный ген или его участок, который необходимо мутагенизировать. В этом случае в образующемся гибриде-гетеродуплексе, одна цепь которого принадлежит вектору без вставки, а другая – вектору со вставкой, происходит выпетливание последовательности вставки в виде одноцепочечного участка ДНК. Обсуждаемый подход к получению статистического набора точковых мутаций с использованием химических мутагенов позволяет легко создавать большое число мутантных молекул ДНК, содержащих одну или несколько мутаций в разных сочетаниях. Последующий отбор мутантов на основе новых биохимических или иных параметров мутантных белков (исчезновение, ослабление или усиление ферментативной активности, появление новой активности или новых иммунологических свойств и т.п.) позволяет идентифицировать остатки аминокислот в исследуемых белках, отвечающие за эти изменения.
Несмотря на удобство введения такого рода мутаций в ДНК in vitro, химический мутагенез накладывает ограничения на спектр возникающих мутаций, так как лишь определенные остатки нуклеотидов ДНК претерпевают обязательные изменения. Поэтому многие мутации никогда не могут быть получены с помощью химических мутагенов. Проблему можно частично решить, используя для репарации одноцепочечных брешей ДНК аналоги нуклеотидов, например N-гидроксицитозинтрифосфат, который в составе ДНК одинаково хорошо спаривается с A и G, или создавая такие условия, при которых репарирующая ДНК-полимераза начинает ошибочно включать в синтезируемую цепь ДНК некомплементарные матрице нуклеотиды. Все перечисленные выше методы локального мутагенеза, осуществляемого in vitro, позволяют, в конечном счете, получать набор случайных мутаций, локализованных на определенном исследуемом участке ДНК. Мутагенизированные молекулы ДНК из одной реакционной пробирки представляют собой сложную смесь, в которой каждая молекула несет несколько независимо возникших мутаций. Для введения мутаций в определенный локус исследуемого гена необходимо проводить трудоемкую процедуру отбора, сопряженную с анализом большого числа мутантов. Подлинную революцию в направленном мутагенезе произвела разработка методов с использованием синтетических олигонуклеотидов.