В живом организме большинство биологических процессов управляется посредством специфических белок-белковых или белково-нуклеиновых взаимодействий. К таким процессам относятся, например регуляция транскрипции генов под действием различных белковых факторов, взаимодействие белковых лигандов с рецепторами на поверхности клеток, а также специфическое связывание антигенов соответствующими антителами. Понимание молекулярных механизмов взаимодействия белковых лигандов с рецепторами имеет большое фундаментальное и прикладное значение. В частности, разработка новых лекарственных препаратов белковой природы обычно начинается с идентификации исходной последовательности аминокислот, обладающей требуемой биологической активностью (так называемая "основная" (lead) последовательность). Однако пептиды с основной последовательностью аминокислот могут обладать и нежелательными биологическими свойствами: низкой активностью, токсичностью, малой стабильностью в организме и т.п.
До появления библиотек пептидов улучшение их биологических свойств осуществляли путем последовательного синтеза большого числа аналогов и проверкой их биологической активности, что требовало больших затрат времени и средств. В последние годы появилась возможность с помощью автоматических синтезаторов создавать за короткое время тысячи различных пептидов. Разработанные методы направленного мутагенеза также позволили резко расширить число белков, получаемых одновременно и последовательно тестируемых на биологическую активность. Однако только недавно разработанные подходы к созданию библиотек пептидов привели к получению миллионов последовательностей аминокислот, требуемых для проведения эффективного скрининга с целью выявления среди них пептидов, максимально удовлетворяющих предъявляемым критериям. Такие библиотеки используются для исследования взаимодействия антител с антигенами, получения новых ингибиторов ферментов и антимикробных агентов, конструирования молекул, обладающих требуемой биологической активностью, или придания новых свойств белкам, например антителам.
По способам получения библиотеки пептидов разделяются на три группы. К первой группе можно отнести библиотеки, полученные с использованием химического синтеза пептидов, в которых индивидуальные пептиды иммобилизованы на микроносителях. При таком подходе после присоединения очередных аминокислот в индивидуальных реакционных смесях к пептидам, иммобилизованным на микроносителях, содержимое всех реакционных смесей объединяют и разделяют на новые порции, которые используют на следующей стадии присоединения новых аминокислотных остатков. После проведения ряда таких этапов оказываются синтезированными пептиды, содержащие последовательности использованных в синтезе аминокислот во всевозможных случайных сочетаниях.
Библиотеки пептидов, иммобилизованных на микроносителях, обладают существенным недостатком: они требуют при скрининге использования очищенных рецепторов, находящихся в растворимой форме. В то же время в большинстве случаев при биологических испытаниях, проводящихся для фундаментальных и фармакологических исследований, чаще всего находят применение рецепторы, ассоциированные с мембранами. По второму способу библиотеки пептидов получают с помощью твердофазного синтеза пептидов, при котором на каждой стадии химического присоединения очередной аминокислоты к растущим пептидным цепям используют эквимолярные смеси всех или некоторых аминокислот-предшественников. На конечной стадии синтеза проводят отделение пептидов от носителя, т.е. перевод их в растворимую форму. Третий подход к конструированию библиотек пептидов, к описанию которого мы сейчас переходим, стал реальным именно благодаря развитию методов генной инженерии. Он прекрасно иллюстрирует возможности таких методов и, несомненно, является крупным достижением в их применении. В этой связи рассмотрим более подробно результаты использования библиотек пептидов в исследовании эпитопов (антигенных детерминант) белков.
Генно-инженерная технология получения гибридных белков позволила разработать эффективный метод наработки коротких пептидов для анализа их биологической активности. Как и в случае клонотек генов, библиотеки пептидов, полученные генно-инженерными методами, представляют собой большой (часто исчерпывающий) набор коротких пептидов. Два недавно сделанных наблюдения позволяют рассматривать библиотеку пептидов одновременно и в качестве библиотеки эпитопов белков. Во-первых, короткие пептиды могут включать все основные остатки аминокислот, играющие главную роль во взаимодействии с антителами, и они в состоянии имитировать крупные антигенные детерминанты белков. Во-вторых, в большинстве случаев нековалентные связи, образуемые между немногими наиболее важными остатками аминокислот белковых лигандов и их рецепторами, вносят основной вклад в общую энергию взаимодействия лиганд–рецептор. С учетом этого любой пептид можно рассматривать как потенциальный лиганд, гаптен или часть антигенной детерминанты более крупных полипептидов, а любую библиотеку пептидов – как библиотеку эпитопов белков или потенциальных лигандов для соответствующих белковых рецепторов.
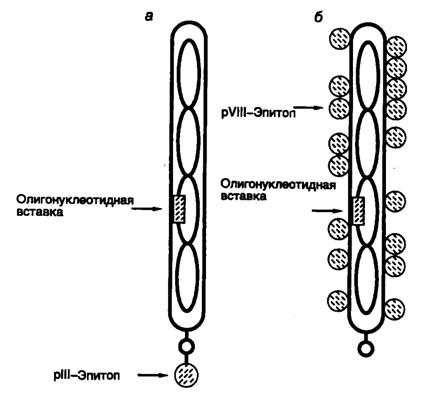
Рис. II.19. Схема экспрессии пептидных эпитопов на поверхности оболочки нитевидных колифагов
Пептидные эпитопы находятся в составе гибридных полипептидных цепей минорного белка pIII (а) или основного белка pVIII вирусной оболочки (б). Стрелки указывают положение кодирующих эпитопы олигонуклеотидных фрагментов в геноме бактериофага, а также положение самих эпитопов. В составе полипептида pIII (а) показана только одна копия эпитопа (на самом деле их число достигает 4–5)
Библиотека пептидов, полученная в результате реализации третьего подхода, в современном виде представляет собой набор десятков или даже сотен миллионов коротких различающихся последовательностей аминокислот, которые экспрессированы на поверхности вирионов бактериофагов в составе их собственных структурных белков. Это становится возможным благодаря введению методами генной инженерии в геном бактериофагов гибридных рекомбинантных генов, кодирующих измененные структурные белки его вирионов. (Данный метод известен под названием фагового дисплея.) В результате экспрессии таких генов образуются гибридные белки, на N- или С-концах которых (см. ниже) присутствуют дополнительные последовательности аминокислот. В наиболее хорошо разработанной системе, позволяющей конструировать библиотеки пептидов генно-инженерными методами, используют небольшой нитевидный колифаг f1 и два его белка: основной и минорный белки оболочки pVIII и pIII. In vivo оба белка синтезируются в виде полипептидных цепей с короткими N-концевыми сигнальными последовательностями, которые отщепляются сигнальной пептидазой во время их созревания после переноса к внутренней части бактериальной мембраны. Зрелые белки встраиваются в оболочку бактериофага в процессе ее сборки. При этом белок pVIII образует основную оболочку бактериофага, тогда как четыре или пять молекул pIII ассоциированы с концевой частью вириона и обеспечивают взаимодействие вирусных частиц с половыми ворсинками клеток E. coli (рис. II.19). Генно-инженерными методами пептиды соединяют с белками – непосредственно с их N-концевыми последовательностями или на небольшом от них расстоянии. Концевые последовательности большинства белков являются более гибкими и, как правило, экспонируются на поверхности глобулы, что позволяет получать гибридные рекомбинантные белки без существенного нарушения их основных свойств, а также делает интегрируемые пептиды доступными для распознавания извне. Кроме того, в таком положении и пространственная структура самих пептидов испытывает меньшее влияние белка-носителя. В ходе экспериментов было установлено, что введение чужеродных пептидов в N-концевую часть белка pIII не оказывает существенного влияния на жизнеспособность и инфекционность фаговых частиц, тогда как соединение пептидов длиной >5 аминокислотных остатков с N-концевой частью белка pVIII нарушает сборку вирионов. Последнее затруднение можно преодолеть доставкой к месту сборки вирионов молекул белка pVIII дикого типа, синтез которых направляется соответствующим геном вируса-помощника. В этом случае оболочка бактериофага будет содержать как измененные белки pVIII, так и полипептиды дикого типа от вируса-помощника.
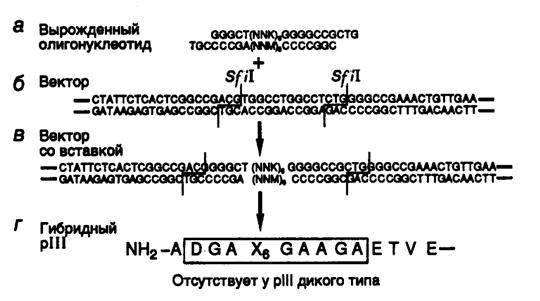
Рис. II.20. Схема конструирования рекомбинантного вирусного генома, содержащего вставки вырожденных олигонуклеотидов, для получения библиотеки эпитопов
Двухцепочечный олигонуклеотид (а), содержащий вырожденные кодоны NNK и те же самые сайты рестрикции в составе линкеров, лигируют с ДНК вектора Fuse5 (б), расщепленного рестриктазой Sfi I, с образованием рекомбинантного генома (в), который направляет синтез гибридного рекомбинантного белка (г), содержащего на N-конце указанную аминокислотную последовательность
При конструировании библиотеки пептидов прежде всего синтезируют два комплементарных друг другу олигонуклеотида, которые после отжига образуют двухцепочечную молекулу, центральная часть которой кодирует собственно пептиды (рис. II.20, а), а выступающие по концам одноцепочечные участки комплементарны "липким" концам вектора, получающимся под действием соответствующей рестриктазы (см. рис. II.20, б).
Для кодирования аминокислот пептидов используют вырожденные кодоны вида NNK или NNS, которые включают все четыре нуклеотида (N) в первом и втором положениях, G или T (K), а также G или С (S) в третьем положении. При таком подходе информация о всех 20 аминокислотах и одном стоп-кодоне заключена в 32 различных кодонах NNK и NNS, а не в 64, как это имеет место в случае природного генетического кода.
В процессе синтеза вырожденных олигонуклеотидов, кодирующих исследуемые пептиды, на каждой стадии используют индивидуальные нуклеотиды для кодонов инвариантных аминокислот, фланкирующих вариабельный участок пептида, а также эквимолярные смеси нуклеотидов для участков, кодирующих случайные последовательности. Образовавшийся в итоге набор вырожденных олигонуклеотидов далее клонируют в виде одноцепочечных фрагментов в соответствующих сайтах гена белка оболочки бактериофага в составе фагового вектора или фазмиды. Альтернативно для такого набора олигонуклеотидов (химически или с помощью ПЦР) синтезируют комплементарные цепи с включением инозина в вариабельные участки, поскольку его остатки, как известно, спариваются с основаниями С и T матрицы, что облегчает образование правильных дуплексов между соответствующими олигонуклеотидами. Образующиеся двухцепочечные олигонуклеотиды в случае необходимости обрабатывают соответствующими рестриктазами и клонируют в фаговом векторе. Итоговые рекомбинантные молекулы (см. рис. II.20, в) ДНК вводят в бактериальные клетки, получая ~109 трансформантов на 1 мг рекомбинантной ДНК, образовавшиеся фаговые частицы размножают в бактериях и после очистки исследуют на присутствие рекомбинантных пептидов (см. рис. II.20, г), способных взаимодействовать с исследуемыми рецепторами в белках их вирионов.
Число индивидуальных фаговых клонов в библиотеке является определяющим для ее использования. К примеру, библиотека, заключающая в себе все возможные гексапептиды, должна содержать 64 млн (206) разных шестичленных аминокислотных последовательностей, кодируемых ~1 млрд (326) различных гексакодонов (32 – число кодонов, с помощью которых можно закодировать любую из 20 аминокислот предложенным выше способом, а именно с использованием кодонов NNK или NNS). Для решения такой задачи должны быть получены очень большие библиотеки, содержащие, по крайней мере, 2·108 – 3·108 индивидуальных, независимых клонов, а величина 109 в настоящее время является верхним пределом для числа индивидуальных клонов библиотеки, которую еще можно практически использовать.
Исходя из этого, можно заключить, что максимальная длина пептидов, включающих в себя все возможные сочетания 20 аминокислот, с которыми возможно работать с помощью библиотек пептидов, составляет 6 аминокислотных остатков. Тем не менее, следует иметь в виду, что библиотека 15-членных пептидов того же размера (2–3·108 клонов) будет содержать больше разнообразных гексапептидов, чем рассмотренная выше библиотека 6-членных пептидов. Кроме того, поскольку лишь ограниченное число аминокислотных остатков в пептиде действительно определяет его биологическую активность, библиотека 15-членных пептидов может оказаться представительнее библиотеки более коротких пептидов с тем же числом клонов.
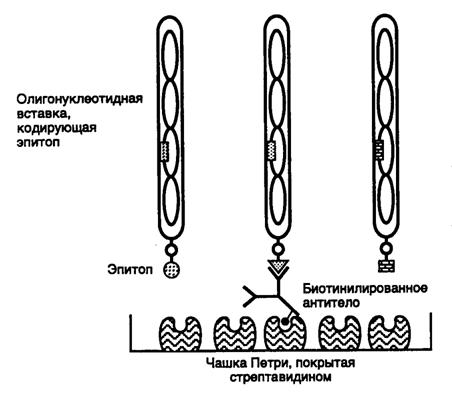
Рис. II.21. Схема отбора фаговых частиц, обладающих требуемыми эпитопами
Показаны три рекомбинантных фаговых частицы, экспрессирующие разные эпитопы в составе pIII. Только эпитоп центральной фаговой частицы распознается молекулой биотинилированного антитела, иммобилизованного на чашке Петри с помощью стрептавидина и использованного для скрининга библиотеки
Для того чтобы выделить из библиотеки пептиды с искомой биологической активностью, применяют различные методы скрининга. В частности, для выделения пептидов, имитирующих определенные эпитопы, используют биотинилированные моноклональные антитела соответствующей специфичности, которые иммобилизуют на твердой подложке с помощью стрептавидина (рис. II.21). Фаговые частицы, экспрессирующие на своей поверхности соответствующие эпитопы, взаимодействуют с антителами и задерживаются подложкой, тогда как другие рекомбинантные фаговые частицы удаляются в процессе промывания. Задержанные на подложке фаговые частицы далее элюируют кислотой, индивидуальные клоны дополнительно размножают в бактериальных клетках и экспрессированные на них эпитопы исследуют по различным критериям. Наличие идентичных или сходных последовательностей нуклеотидов среди клонированных последовательностей свидетельствует о специфичности процесса очистки. Индивидуальные клоны затем охарактеризовывают другими, в частности иммуноферментными методами. На заключительной стадии исследования осуществляют синтез выделенных пептидов и их всестороннее изучение в очищенном состоянии.
В настоящее время имеются данные о некоторых работах, проведенных с использованием пептидных библиотек. В одном из таких исследований из библиотеки были выделены пептиды, последовательность аминокислот в которых резко отличалась от последовательности аминокислот истинного эпитопа исследуемого антигена. Тем не менее, такой пептид прочно связывался со специфическими антителами и конкурировал за связывание с природным антигеном. Это позволило сделать вывод о возможности существования мимотопов – коротких пептидов, имитирующих природные эпитопы, последовательности аминокислот которых существенно различаются между собой. Удалось установить канонические последовательности аминокислот пептидов, имитирующих эпитопы природных белков, а среди них идентифицировать аминокислотные остатки, играющие ключевую роль во взаимодействии антиген–антитело.
Одним из многообещающих приложений библиотек пептидов является идентификация пептидных лигандов, имитирующих "структурные" эпитопы, образующиеся на поверхности белковых глобул в результате сворачивания их полипептидных цепей, что сопровождается пространственным сближением аминокислотных остатков, расположенных в полипептидной цепи на значительном расстоянии друг от друга. С помощью пептидных библиотек возможна идентификация пептидных аналогов различных эпитопов небелковой природы. По-видимому, в ближайшем будущем возможно использование пептидных библиотек для получения новых лекарственных препаратов, создания диагностических средств и производства эффективных вакцин. В области конструирования новых лекарственных препаратов усилия исследователей могли бы быть направлены на создание пептидных лигандов, специфически взаимодействующих с рецепторами, представляющими медико-биологический интерес. Знание структуры таких лигандов позволило бы упростить получение на этой основе лекарственных препаратов небелковой природы.
Библиотеки пептидов и эпитопов найдут свое применение и в исследованиях механизмов гуморального иммунного ответа, а также заболеваний иммунной системы. В частности, большинство аутоиммунных заболеваний сопровождается образованием аутоантител против антигенов собственного организма. Эти антитела во многих случаях служат специфическими маркерами того или иного аутоиммунного заболевания. С использованием библиотеки эпитопов, в принципе, можно получить пептидные маркеры, с помощью которых было бы возможно следить за специфичностью аутоантител во время развития патологического процесса как в индивидуальном организме, так и в группе пациентов и, кроме того, определять специфичность аутоантител при заболеваниях неизвестной этиологии.
Библиотеки пептидов и эпитопов потенциально могут быть использованы также для скрининга иммунных сывороток с целью выявления пептидов, специфически взаимодействующих с защитными антителами. Такие пептиды будут имитировать антигенные детерминанты патогенных организмов и служить мишенями для защитных антител организма. Это позволит использовать подобные пептиды для вакцинации пациентов, у которых отсутствуют антитела против соответствующих патогенов. Изучение эпитопов с помощью библиотек пептидов является частным случаем одного из многочисленных направлений их использования в прикладных и фундаментальных исследованиях взаимодействия лигандов и рецепторов. Дальнейшее усовершенствование этого подхода должно способствовать созданию новых лекарственных препаратов на основе коротких пептидов и быть полезным в фундаментальных исследованиях механизмов белок-белковых взаимодействий.