На определенном этапе онтогенеза верхушечная меристема побега начинает вместо листьев, междоузлий формировать цветки или соцветия. Однако она должна быть индуцирована для перехода к генеративному развитию. Внешними индукторами этого процесса являются температура, свет, продолжительность дня и ночи, вода, элементы минерального питания.
Пониженные температуры (яровизация). У некоторых растений способность к заложению цветков, т.е. переход к этапу зрелости, появляется лишь после действия на них пониженных температур в течение определенного времени. Уже в старину люди знали, что злаки делятся на двулетние и однолетние – озимые и яровые.
Все растения можно разделить на три группы:
1. растения, которые не зацветают без действия пониженных температур (озимые, двулетники, некоторые многолетники);
2. растения, которые ускоряют зацветание после действия на них пониженных температур, но это действие не обязательно (салат, шпинат);
3. растения, которым не нужны пониженные температуры для перехода к цветению (яровые).
Озимые рожь, пшеница, ячмень при весеннем посеве интенсивно растут, кустятся, но в отличие от яровых форм у них в первый год не закладываются цветки. Действию пониженных температур озимые подвергаются в естественных условиях осенью и весной, поэтому на следующий год после посева нормально выколашиваются. Свойство яровых цвести в первый год жизни, а озимых во второй – генетически запрограммировано. Свойство озимых однолетних и двулетних растений ускорять переход к заложению цветков после действия на них пониженных температур в течение определенного времени назвали яровизацией.
У большинства растений во время действия яровизирующих температур цветки не закладываются, что обычно происходит после наступления благоприятных для роста более высоких температур. Пониженные температуры действуют как индуктор. Однако у некоторых растений заложение цветков происходит только при пониженных температурах. Рецептором, воспринимающим действия пониженных температур, является точка роста.
Под влиянием пониженных температур в клетках меристемы происходят изменения, подготавливающие заложение цветков при наступлении благоприятных условий: увеличивается количество РНК, активируются гидролитические ферменты, изоэлектрическая точка белков сдвигается в кислую сторону, усиливаются окислительные процессы.
Растениям разных видов для подготовки к заложению цветков необходимы различные пониженные температуры. Обычно наиболее эффективны температуры порядка + 1 – +7 °С. Продолжительность периода охлаждения и значения эффективных температур зависят от вида и даже разновидности (сорта) растений. У большинства растений этот период составляет 1-3 месяца, у других – от нескольких дней до двух недель. Если холодный период слишком короток или прерывается повышением температур выше 15 °С, то яровизация замедляется или совсем не происходит.
Яровизация является приспособлением к неблагоприятным условиям осеннего и весеннего периодов, когда бывают заморозки; цветки закладываются только после установления стабильных повышенных температур.
Яровизационные изменения постепенно накапливаются и передаются при делении меристематических клеток новым клеткам. Боковые меристемы, образовавшиеся из яровизированной меристемы, сохраняют эти изменения. Однако семена озимых однолетних и двулетних культур, полученные с яровизированных растений, нуждаются в новой яровизации. Предполагают, что потеря яровизационных изменений происходит во время мейоза при образовании мега- и микроспор.
Для прохождения яровизации необходимы, прежде всего, кислород и вода. Если наклюнувшиеся семена выдерживать в анаэробных условиях, например в атмосфере азота или водорода, то яровизация не происходит. Следовательно, происходящие процессы связаны с дыханием. Для прохождения яровизационных процессов клетки должны содержать довольно много воды, например у злаков – около 35 %. Для яровизации необходимы также сахара.
Яровизационные изменения обратимы. Под действием повышенных температур, равных +25 – +40 °С, возможна разъяровизация: готовность меристемы к заложению цветков теряется.
Продолжительность дня и ночи (фотопериодизм). Зависимость зацветания растений от соотношения светлой и темной частей суток называют фотопериодизмом, а соотношение длины дня и ночи – фотопериодом.
При изучении этого явления оказалось, что длина дня действует на зацветание разных растений неодинаково. Сначала были обнаружены три типа растений: короткодневные, длиннодневные и нейтральные.
Короткодневные – это растения, которые зацветают при длине дня 8-12 ч в сутки; находясь все время в более длинном дне, они только интенсивно растут. Из культурных растений короткодневными являются рис, кукуруза, просо, соя, сахарный тростник, хлопчатник, сорго.
Длиннодневные – это растения, которые зацветают при продолжительности дня не менее 16-18 ч/сут; на коротком дне они приобретают розеточную форму. При непрерывном освещении эти растения еще сильнее ускоряют зацветание, поэтому на Севере у них короткий вегетационный период. К этому типу растений относятся пшеница, ячмень, овес, горчица, свекла, лен, шпинат, клевер.
Нейтральные – это растения, зацветающие при любой продолжительности дня. К ним относятся фасоль, томаты. Разные сорта одного вида, например табака, могут быть длиннодневными и короткодневными.
В последние годы выделены еще длинно-короткодневные растения и коротко-длиннодневные. Первые для перехода из вегетативного в генеративное состояние должны сначала развиваться при длинном, а затем – при коротком дне. Такие растения цветут осенью. Коротко-длиннодневным растениям для зацветания нужен сначала короткий, а потом длинный день. В каждой из этих групп есть растения, обязательно требующие для зацветания соответствующего фотопериода (облигатные виды), и растения, только ускоряющие зацветание при нужной длине дня (необлигатные виды).
Фотопериодическое воздействие вызывает свет малой интенсивности, на порядок меньше того, при котором идет фотосинтез. Даже яркий лунный свет может быть причиной незацветания короткодневных растений. Это доказывает относительную независимость фотопериодизма от фотосинтеза.
Продолжительность дня или ночи воспринимает листовая пластинка, которая воспринимает длину дня и ночи, но изменения, в результате которых начинается заложение цветков, происходят в меристеме. На фотопериодическое воздействие отвечает ближайшая к листу точка роста. Чувствительность листовой пластинки зависит от возраста листа. Наибольшая чувствительность совпадает по времени с достижением листом максимального размера. Очень молодые или старые листья менее чувствительны, чем развернувшиеся.
Продолжительность дня и ночи листья воспринимают с помощью фитохрома. Так как меристематические ткани характеризуются наиболее высоким содержанием фитохрома, то предполагают, что верхушки побегов также способны реагировать на изменение длины дня.
Механизм фотопериодического действия можно представить следующим образом. Фитохром поглощает свет, активизируется и вызывает синтез гормонов в листе. В меристеме под влиянием притекающих туда гормонов происходят изменения, приводящие к заложению цветка. Чтобы гормоны оказали влияние на меристематические клетки, они должны быть компетентны.
Длинно- или короткодневность растений зависит от географического происхождения вида или сорта. Как правило, тропические растения – короткодневные, растения умеренной зоны и приполярной – длиннодневные. Когда после зимы постепенно дни становятся длиннее и опасность заморозков исчезает, на длинном дне закладываются цветки. В тропических и субтропических странах период более длинных дней часто совпадает с засухой или ливнями. Оба явления неблагоприятны для опыления растений и созревания плодов, вследствие чего в этих широтах, кроме нейтральных, распространены короткодневные растения. Таким образом, длиннодневность выработалась у растений в связи с перезимовкой, короткодневность – в связи с периодическими засухами или тропическими ливнями.
Неблагоприятные факторы: засуха, чередующиеся пониженные и повышенные температуры – в разные годы приходятся на различные календарные даты, а продолжительность определенного дня месяца каждый год одинакова, так как зависит от географической широты местности. Поэтому длина дня служит надежным ориентиром для изменения сезонных ритмов в росте и развитии. Уменьшение длины дня в умеренной зоне является для растений сигналом приближения периода пониженных температур, а удлинение дня в тропиках и субтропиках – сигналом наступления засухи.
Следовательно, фотопериодизм, как и яровизация, – это адаптация к неблагоприятным условиям среды, позволяющая растениям зацветать в наиболее благоприятное время года.
Короткий или длинный день нужен растению не всю жизнь, а лишь определенное время, после чего оно зацветает при любой длине дня. Растения отличаются друг от друга по числу суток с определенной длиной дня, необходимых для перехода к цветению. Таким образом, фотопериод выступает как индуктор (сигнал), в результате которого наступает цветение. Можно дать растению необходимый фотопериод в самом начале онтогенеза (ювенильный этап), а потом выращивать его все время при любой длине дня, и растение зацветет. Это явление получило название фотопериодической индукции.
Отсюда можно сделать важный вывод: при интродукции южных короткодневных растений на север, в условиях длинного дня, необязательно выращивать их до цветения в условиях короткого дня, достаточно лишь дать им длинную ночь в течение определенного числа суток.
Кроме заложения цветков, фотопериод влияет и на другие процессы, например, на листопад, формирование вегетативных органов. От фотопериода зависит также формирование колючек, шипов, усов, прямостоячих, стелющихся или вьющихся побегов, суккулентных или мезофитных листьев.
Элементы минерального питания. Еще до открытия фотопериодизма и яровизации наблюдения за зацветанием растений показали, что у некоторых видов, например у плодовых деревьев, цветение и бурный рост являются антагонистами. Факторы, тормозящие рост: засуха, обрезка, кольцевание – стимулируют цветение. Наоборот, повышенное азотное питание, усиливающее рост, тормозит цветение.
У некоторых видов растений инициация цветения находится в прямой зависимости от наличия железа и меди. Низкое содержание ионов меди снижает потребность в определенной длине дня. Короткодневное растение ряска зацветает в этом случае на длинном дне. Так же действует и железо. Механизм действия этих элементов неизвестен.
Водный дефицит играет важную роль в инициации цветения у большинства однолетних растений. Они зацветают быстрее в условиях засухи. Водный стресс вызывает гидролиз крахмала и белка, что приводит к значительному повышению содержания растворимых углеводов и аминокислот, особенно пролина. Двулетние и многолетние растения, наоборот, задерживают цветение в этих условиях.
Таким образом, у растений существуют три типа контроля за зацветанием: возрастной, яровизационный и фотопериодический.
Гормоны цветения. М.Х. Чайлахян выдвинул гипотезу, согласно которой цветение вызывают два гормона: один из них – гиббереллин, а другой – неизвестный гормон, названный антезином. Процесс зацветания проходит в две фазы: на первой фазе образуется цветочный стебель (цветоножка или ось соцветия и цветоножки), а на второй – сами цветки. У длиннодневных растений критической является первая фаза зацветания, которая зависит от присутствия гиббереллина, синтезирующегося в листьях на длинном дне. Антезин у этих растений всегда имеется в достаточном количестве.
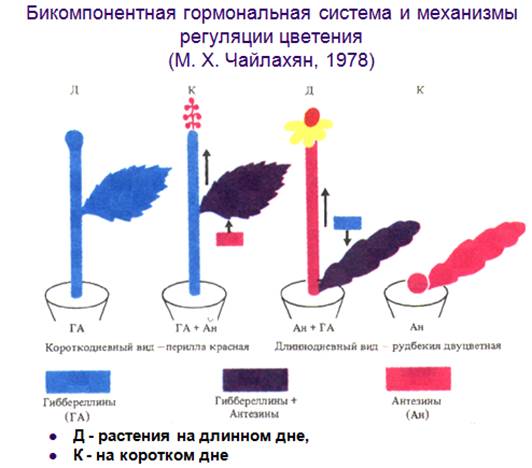
Короткодневные растения, наоборот, при любой продолжительности дня содержат много гиббереллинов, поэтому опрыскивание их гиббереллином не вызывает зацветания: им не хватает антезинов, образующихся на коротком дне. Итак, в листьях при благоприятной продолжительности дня образуется недостающий гормон.
Нейтральные растения цветут при любой длине дня, так как у них есть и гиббереллины, и антезины; образование цветков у них зависит только от возраста растения.
Оба гормона транспортируются в точки роста, расположенные ближе всего к листовой пластинке. В точках роста происходят соответствующие изменения: усиливаются репликация ДНК, синтез мРНК и специфических белков, что приводит в конечном счете к формированию цветков. Для того чтобы гормоны оказали влияние, меристематические клетки должны быть компетентными. Эта компетенция возникает у клеток в определенном возрасте, разном у различных растений.
Формирование мужских и женских цветков. У растений чаще образуются обоеполые (гермафродитные) цветки, реже однополые – мужские или женские. Поскольку клетки каждого растения тотипотентны, то потенциально они могут обеспечить формирование цветков обоих полов. Какие цветки образуются на растении, зависит от генотипа.
Короткий день или уменьшение интенсивности света вызывает массовое появление на мужских растениях обоеполых или даже женских цветков. Длинный день действует в противоположном направлении. Синий свет усиливает женскую сексуализацию, а красный – мужскую. Регуляция пола у раздельнополых растений контролируется фитохромом.
В системе, регулирующей сексуализацию цветков, есть главные факторы: гормоны цитокинины и гиббереллины. В женских цветках цитокининов содержится значительно больше, чем в мужских. Цитокинины, образующиеся в корнях, транспортируются в верхушки стеблей и индуцируют образование в меристеме женских (пестичных) цветков. Гиббереллины, синтезируемые в листьях, транспортируясь в верхушечные меристемы, вызывают образование мужских (тычиночных) цветков.
На формирование мужских и женских цветков влияют и внешние условия. Все факторы, способствующие корнеобразованию, усиливают формирование женских цветков, так как в сильно развитой корневой системе образуется большое количество цитокининов; наоборот, факторы, вызывающие усиление роста побегов, в частности листьев, индуцируют образование мужских цветков, так как в хорошо развитых листьях увеличивается синтез гиббереллинов.
На заложение мужских или женских цветков влияют условия минерального питания. Избыток солей кальция в почве способствует развитию большого количества мужских цветков, а внесение фосфорных удобрений приводит к увеличению числа женских цветков.
Повышенная влажность почвы и воздуха в период заложения цветков – необходимое условие для образования женских цветков, а понижение влажности способствует формированию мужских цветков. Окись углерода, этилен, ацетилен стимулируют образование женских цветков. Низкие температуры способствуют формированию женских цветков, а высокие – мужских.
Быстрый рост растений стимулирует образование мужских цветков, а медленный – женских. Итак, короткий день, синий свет, СО, высокий уровень азота, высокая влажность и низкие температуры, способствующие корнеобразованию, усиливают формирование женских цветков. Наоборот, такие факторы, как длинный день, красный свет, высокий уровень кальция, высокие температуры и относительно низкая влажность, вызывающие усиление роста побегов, индуцируют образование мужских цветков.