Ритмические сокращения сердца обеспечиваются последовательным прохождением электрического импульса по проводящей системе сердца. В норме электрический импульс, вызывающий сердечное сокращение, вырабатывается в синоатриальном узле, распространяется в мышце предсердия через межклеточные контактные мостики - вставочные диски, которые обеспечивают непрерывность распространения импульса между клетками и достигают А-В узла.
В связи с тем, что предсердно-желудочковые клапаны окружает фиброзная ткань, распространение электрических импульсов от предсердий к желудочкам возможно только через А-В узел. Как только электрический импульс достигает А-В узла, происходит задержка его дальнейшего проведения на 0,1 секунды. Эта задержка объясняется проведением импульса через А-В узел по медленным каналам.
Пауза в проведении импульса полезна:
- т.к. она дает предсердиям время для их сокращения до начала возбуждения и сокращения желудочков;
- задержка позволяет А-В узлу выполнить функцию привратника, препятствуя проведению слишком частых импульсов от предсердий к желудочкам при предсердных тахикардиях.
Выйдя из А-В узла, сердечный потенциал действия распространяется по системе Гиса-Пуркинье к основной массе клеток миокарда, что обеспечивает координированное сокращение кардиомиоцитов.
В основе проведения электрических импульсов по проводящей системе сердца, сокращения миокарда лежат процессы электрической стимуляции клеток с формированием потенциала возбуждения, который образуется за счет ионных токов через специальные каналы сарколеммы; то есть деполяризации и реполяризации клеток.
В состоянии покоя внутри клетки концентрация катионов калия в 30-35 раз выше, чем в межклеточной жидкости, а концентрация натрия в 10-20 раз меньше. В состоянии покоя открыты лишь калиевые каналы, по которым К+ выходит из клетки, а поскольку внутри клетки они связаны с белковыми комплексами, имеющими отрицательный заряд, то, при выходе калия из клетки, внутренняя мембрана приобретает отрицательный заряд, а наружные - положительный заряд. Равновесие противоположных зарядов внешней и внутренней сторон мембраны клетки называется статической поляризацией. Разность потенциалов между внутренней и внешней мембраной клетки составляет - 90 МВ и называется потенциалом покоя. Любое воздействие, которое делает заряд мембраны еще менее отрицательным (например, образовавшийся спонтанно импульс С-А узла) приводит к открытию натриевых каналов и входу Na+ внутрь клетки. При этом внутренняя поверхность мембраны становится положительно заряженной, а внешняя отрицательно - происходит деполяризация клеточной мембраны. Электрический потенциал, возникающий в момент возбуждения клетки, называют потенциал действия. После фазы деполяризации клеточной мембраны следует реполяризация.
При реполяризации К+ выходит из клетки, Са входит в клетку, и внутренняя сторона клеточной мембраны вновь приобретает отрицательный заряд, а внешняя положительный. Фаза реполяризации переходит в фазу покоя - включаются АТФ - зависимые (энергопотребляемые) Na+/К+ и кальциевые насосы, восстанавливающие нормальные трансмембранные градиенты ионов внутри и вне клетки.
Некоторые сердечные клетки не нуждаются во внешних стимулах, а сами способны инициировать деполяризацию (возбуждение) - это пейсмекерные клетки. Они обладают автоматизмом - способностью к спонтанной деполяризации. Наиболее широко они представлены в С-А узле, далее А-В узле и системе Гиса-Пуркинье - соответственно водители ритма I - ого, II - ого и III - го порядка.
Потенциал покоя представлен фазой 4 потенциала действия. После деполяризации проникновение Na+ внутрь клетки приводит к быстрому наступлению фазы 0; выход тока калия наружу обусловливает частичную реполяризацию во время фазы 1; медленное проникновение Са++ внутрь (и относительно медленный выход К+ из клетки) приводит к временному выравниванию напряжения (плато на рис.8) - фаза 2; завершающая быстрая реполяризация обусловлена в основном выходом К+ во время фазы 3.
При входе кальция внутрь кардиомиоцита во время возбуждения становится возможным взаимодействие актина и миозина - сократительных белков, что приводит к сокращению клеток и миокарда в целом. Для полноценного расслабления миокарда Са закачивается в саркоплазматический ретикулум с помощью «Са+/насоса» и удаляется из клетки.
Во время абсолютного рефрактерного периода (АРП) клетка нечувствительна к стимуляции. Эффективный рефрактерный период помимо АРП включает короткий период, в течение которого стимуляция вызывает локальную деполяризацию, которая не способна распространяться. Во время относительного рефрактерного периода стимуляция вызывает слабый потенциал действия (ПД), который распространяется дальше, но более медленно, чем обычный.

Рис. 8. Схема потенциала действия (ПД) миоцита и ионные токи для Na+, Ca++, и К+.
В периоде сверхнормальной возбудимости более слабые стимулы, чем в норме, могут вызывать ПД.
Рефрактерный период - период, во время которого клетки не восприимчивы к повторным стимулам. Выделяют абсолютно рефрактерный период, когда клетки полностью нечувствительны к новым стимулам и относительно рефрактерный период, когда возникновение потенциала действия (возбуждение, ответ) клетки возможно под воздействием более сильного раздражителя.
После относительно - рефрактерного периода выделяют период сверх нормальной возбудимости, в которой раздражители даже меньшей силы способны вызвать ответ. Это так называемый уязвимый период, когда могут возникать ранние экстрасистолы и другие жизнеопасные нарушения сердечного ритма.
Рефрактерный период клеток предсердия короче, чем клеток миокарда желудочков, поэтому ритм предсердий может значительно превышать частоты сокращений желудочков.
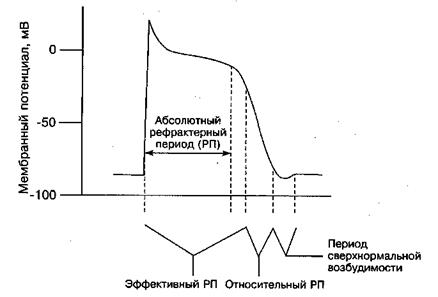
Рис. 9. Рефрактерные периоды (РП) кардиомиоцита.