Координации- устойчивые взаимозависимости органов и систем, проявляющиеся в филогенезе.
Три группы координаций (по механизмам преобразований биологических структур):
· Биологические координации наблюдаются между структурами, непосредственно не связанными ни по функциям, ни по месту положения. Связующее звено между ними- адаптация к определенным условиям обитания. У большинства эндопаразитов сильны развиты половая система и органы прикрепления к телу хозяина, но при этом недоразвиты органы чувств и опорно-двигательный аппарат. Все позвоночные, имеющие матку, обладают также совершенной системой терморегуляции и выкармливают своё потомство молоком.
· Динамические координации выражается во взаимном соответствии структур, связанных функционально. Тесные динамические координации имеются между органами кровеносной и дыхательной систем. Животные, дышащие легкими, имеют 3-х или 4-х камерное сердце и два круга кровообращения.
· Топографические координации проявляются между структурами, связанными друг с другом пространственно. Например, для каждого типа животного царства характерен своеобразный общий план строения, выражающийся в определенном взаимном расположении основных органов и систем. У всех представителей типа хордовых на спинной стороне тела развивается нервная трубка, под ней лежат хорда, пищеварительная трубка и брюшной кровеносный сосуд, а по бокам тела-производные мезодермы.
Все типы координаций имеют высокую степень устойчивости.
Ведущая филогенетическая координации – биологические, поскольку именно их в первую очередь контролирует естественный отбор.
Онтогенетическая корреляция – сложная система связей, обусловливающая развитие организма как единого целого.
· Геномные корреляции. Основным механизмом является зависимость между развитием признаков определяемая действием связанных генов (множественное действие гена, пенетрантность, множественный аллелизм). Например: сочетание безрогости и короткошерстности у коз, мутации у плодовой мухи: редукция крыльев и одновременное укорочение задней пары ног.
· Морфогенетические возникают на ранних стадиях эмбрионального развития, когда функциональные связи между частями зародыша отсутствуют. При этом зачатки, образующиеся раньше, определяют развитие и характер изменения зачатков, возникающих позже. В основе таких корреляций лежит эмбриональная индукция. Пример: соотношение в величине, форме и расположении отдельных органов и частей в организме.
· Эргонтические корреляции проявляются на более поздних стадиях онтогенеза в растущем и взрослом организме. Они обусловлены функциональной взаимозависимостью различных органов или частей организма. Например: корреляции между развитием периферических органов, нервов и нервных центров в ЦНС; зависимость между степенью развития мышц и костного выступа; ход развития половых желез и вторичных половых признаков или некоторых эндокринных желез и скелета.
Система корреляций представляет собой главный фактор нормального онтогенеза, обусловливающий согласованное течение формообразовательных процессов. Корреляции могут оказывать на онтогенез частное или общее влияние.
Корреляции общего характера, сохраняющие свое полезное значение при широком круге условий, даже при глубоких преобразованиях организма, сохраняются естественным отбором. Примером может служить взаимозависимость между степенью развития мышц и нервным центром, контролирующим их функцию. А также степенью развития органа и кровеносных сосудов, его снабжающих. Такая корреляция полезна и она сохраняется в процессе эволюции.
В результате, в ходе исторического развития происходит медленное, но неуклонное накопление корреляций общего значения, что обусловливает преемственность форм, выражающуюся вповторении предковых черт организации.
81.Общий план строения хордовых. Узловые моменты в прогрессивной эволюции хордовых и их рекапитуляции в онтогенезе человека.
Хордовые представляют собой наиболее высокоорганизованный тип в животном мире. Их общими чертами являются следующие.
1. Внутренний осевой скелет представлен хордой, которая присутствует в эмбриогенезе у всех представителей типа, а у высших дополняется, а затем и замещается позвоночником.
2. Над хордой располагается центральная нервная система в виде нервной трубки с полостью — невроцелем.
3. В боковых стенках глотки находятся жаберные щели, соединяющие ее полость с внешней средой. У рыб и некоторых земноводных они сохраняются в течение всей жизни, у высших хордовых — только в эмбриональномпериоде.
4. Тело построено метамерно. У низших хордовых и у зародышей высших сегментация распростраянется на все системы органов, у высших ярко выражена только в эмбриональном периоде. Позже частично сохраняется только в опорно- двигательном аппарате, нервной и кровеносной системах.
5. Органами поддержания равновесия и движения являются конечности, причем у низших хордовых большее значение имеют непарные, а у высших — парные.
Узловые моменты в прогрессивной эволюции хордовых.
I —появление хрящевого скелета, дифференцировка центральной нервной системы на головной и спинной мозг
II— появление челюстей
III —появление парных конечностей наземного типа и легких
IV— преодоление барьера влажности
V — теплокровность
VI— внутриутробное развитие
82. Эволюция общего плана строения кровеносной системы хордовых и сердца позвоночных. Прогрессивные направления и способы филогенетический преобразований. Врожденные пороки развития у человека.
• Ланцетник:
- Кровеносная система замкнутая, 1 круг кровообращения;
- Нет сердца, функция – пульсирующая брюшная аорта, от которой отходят 100-150 пар Жаберных артерий (не формируют капилляры), расположенных в стенке глотки;
- Артериальная кровь из ЖА àКорни спинной аорты;
§ Передние ветви – Сонные артерии;
§ Задние – сливаются в Спинную аорту на уровне заднего конца глотки;
- Спинная аорта разветвляется на многочисленные артерии, несущие артериальную кровь к органам.
- После обмена в тканях венозная кровь àПарные передние и задние Кардинальные веныàОбщие Кардинальные вены (Кювьеровы протоки) àБрюшная аорта;
- Венозная кровь от кишечника àПодкишечная вена àВоротная система печени àпеченочная вена àБрюшная аорта;
· Рыбы:
- Часть Брюшной аорты из-за интенсификации её функции àДвухкамерное сердце (имеет Венозный синус, предсердие, желудочек, артериальный конус);
- От сердца отходит Брюшная аорта, несёт венозную кровь к жабрам;
- Жаберные артерии олигомеризуются до 4-5 пар, распадаются на капилляры;
- В остальном – соответствует оной у ланцетника;
· Земноводные:
- Замена жаберного дыхания на лёгочноеà2 круга кровообращения;
- Сердце трёхкамерное, сохраняется венозный синус и артериальный конус;
- От сердца отходит Общий артериальный ствол, последовательно ветвящийся на:
§ Кожно-лёгочные артерии;
§ Дуги аорты;
§ Сонные артерии;
- Венозная кровь – из Правого предсердия в Желудочек;
- Артериальная кровь – из левого предсердия в желудочек;
- Полного смешения крови нет, ибо есть мышечные перекладины;
§ Из Правой половины желудочка àвенозная кровь в Кожно-лёгочные артерии;
§ Из центральной части àСмешанная кровь в дуги аорты;
§ Из левой части àАртериальная кровь в Сонные артерии;
- Дуги аорты позади сердца сливаются в Спинную аорту, распадающуюся на многочисленные артерии, питающие органы;
- Задние кардинальные вены – редуцируются, собирают кровь только с боковых поверхностей тела, их замещает функционально Задняя полая вена (в неё впадает печеночная вена);
- Передние кардинальные вены теперь называются Яремными венами;
- Кювьеровы протоки = Передние полые вены;
· Рептилии:
- Трёхкамерное сердце, нет венозного синуса и артериального конуса;
- Дифференцировка общего артериального ствола:
§ Правая дуга аорты – артериальная кровь;
Ø От ПД отходят обе сонные и подключичные артерии;
§ Левая дуга аорты – смешанная кровь;
§ Лёгочный ствол (делится на 2 лёгочные артерии) – венозная кровь;
§ Обе дуги позади сердца сливаются в Спинную аорту;
· Млекопитающие:
- Четырёхкамерное сердце;
- Полное разделение венозной и артериальной крови àпредпосылка к теплокровности;
- От Левого желудочка отходит Левая дуга аорты, которая продолжается в Спинную Аорту àартерии к органам;
§ Точнее, отходят обе Дуги, но правая редуцирована до Плече-Головного ствола, который делится на правые Сонную и Подключичную артерии);
§ Левые отходят от Левой дуги;
- От Правого желудочка отходит лёгочный ствол, который делится на 2 лёгочные артерии;
· Асимметрия основных венозных сосудов:
- Правая передняя полая вена принимает в себя Безымянную, образовавшуюся при слиянии Ярёмной и Левой подключичной;
- Рудименты Задних кардинальных вен – Непарные (позвоночные) вены, соединяются анастомозом:
§ Правая – непарная;
Ø Впадает в Правую переднюю полую вену;
§ Левая – полунепарная;
Направленияи способы преобразований:
o Гомология сердца – общность происхождения и плана строения сердца;
o Гетеротопия сердца – изменение положения;
Ø Глоточная область – Рыбы;
Ø Грудная – наземные позвоночные;
o Гетерохрония закладки сердца – более ранняя закладка у Высших позвоночных;
o Олигомеризация – редукция 1, 2 и 5 Жаберных дуг;
o Смена функций – преобразование жаберных дуг:
Ø 3 – Сонные артерии;
Ø 4 – дуги аорты;
Ø 6 – Лёгочные артерии;
o Смена функций – формирование Плече-головного ствола из сильно редуцированной Правой дуги аорты;
o Субституция – замещение Задних кардинальных вен на Задние полые (у земноводных);
Ø Одновременное Ослабление функций Задних кардинальных, которые превращаются в Непарные вены, собирающие кровь с боковых поверхностей тела;
o Анаболия – формирование капиллярной сети жаберных артерий у рыбёх;
o Девиация – редукция и дифференцировка жаберных дуг;
Пороки развития:
- Персистирование дуг аорты – обе дуги срастаются (как у Земноводных и Рептилий) позади пищевода и трахеи, образуя нисходящую часть спинной аорты;
- Персистирование артериального (боталлова) протока – часть корня спинной аорты меж 4 и 6 парами артерий слева. Сброс артериальной крови из БКК в МКК;
- Персистирование первичного эмбрионального ствола – из сердца выходит только один сосуд;
- Транспозиция сосудов – аорта отходит от Правого желудочка, а Лёгочный ствол – от левого;
- Двухкамерное сердце и шейная эктопия сердца – рыбки;
- Дефект межпредсердной перегородки;
- Дефект межжелудочковой перегородки;
- Развитие «аортального ствола» - есть обе дуги аорты;
83. Эволюция артериальных жаберных дуг позвоночных. Прогрессивные направления и способы филогенетических преобразований. Врожденные пороки развития у человека.
Рыбы. Во время эмбриогенеза закладываются 5-7 пар жаберных артерий, затем 1, 2 и 7-я - редуцируются, а 3-6 пары остаются функционировать. У земноводных закладываются во время эмбриогенеза 6-7 пар жаберных артерий: 1, 2, 5 и 7 – редуцируются, из 3-ей развиваются сонные артерии, из 4-ой – дуги аорты, из 6-ой – кожно-легочные артерии. У рептилий закладываются 6 пар жаберных артерий. Они преобразуются в те же сосуды, что и у земноводных (6 пара – в легочные
артерии). У млекопитающих закладываются 6 пар жаберных артерий, затем 1-я и 2-я пары
редуцируются; 3-я дает сонные артерии; 4-я правая редуцируется, а левая преобразуется в левую дугу аорты; 5-я – редуцируется; 6-я дает легочные артерии.
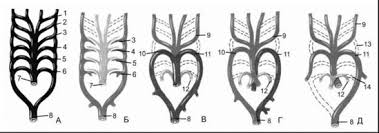
Рис. Развитие артериальных дуг позвоночных животных. А – зародыш позвоночных, Б – рыба, В – бесхвостая амфибия, Г – рептилия, Д – млекопитающее. 1-6 – артериальные (жаберные) дуги, 7 – брюшная аорта, 8 – спинная аорта; 9 – сонные артерии, 10 – правая дуга аорты, 11 – левая дуга аорты, 12 – легочные артерии, 13 – сонный проток, 14 – боталлов проток.
Преобразование 3 артериальных жаберных дуг в сонные артерии, 4 – в дуги аорты, 6 – в легочные артерии – смена функций. Пример олигомеризации – редукция 1, 2 и 5 пар артериальных жаберных дуг.
Пороки развития. Неразделенный аортальный ствол, транспозиция сосудов, отходящих от сердца, сужение аортального или легочного ствола, персистирование артериального (боталлова) протока, развитие «аортального кольца» (наличие обеих дуг аорты), развитие правой дуги аорты и правого корня спинной аорты вместо левого.
84. Место человека в системе животного мира. Методы изучения эволюции человека.
Место человека в системе животного мира:
Человек как биологический вид относится к:
· Царству Животных (Animalia)
· Типу Хордовых (Chordata)
· Подтипу Позвоночных (Vertebrata)
· Классу Млекопитающих (Mammalia)
· Отряду Приматов (Primates)
Основные этапы антропогенеза:
| Временные границы | Этапы антропогенеза | Характерные черты развития |
| 40 тыс. лет | Стадия неоантропа. Вид Человек разумный — Homo Sapiens | Формирование облика современного человека. Возникновение общества. Одомашнивание растений и животных |
| 200-500 тыс. лет | Стадяя палеоантропа. Вид Человек неандертальский — Homo neanderthalensis | Объем головного мозга 1200-1400 см2. Высокая культура изготовления орудий труда. Совершенствование речи и племенных отношений |
| 1-1,3 млн. лет | Стадия архантропа (питекантроп — о.Ява; синантроп — Китай, атлантроп — Африка, гейдельбергский человек — Европа). Вид Человек прямоходящий. Вид Человек умелый. | Объем мозга 800-1200 см2. Формирование речи. Овладение огнем. Переходная стадия к формированию типа современного человека. Объем мозга 500-800 см2. Изготовление первых орудий труда (галечная культура) |
| 2-2,5 млн. лет | ||
| 9 млн. лет | Стадия протантропа. Австралопитеки — предшественники людей. | Переходная форма обезьяны к человеку. Прямоходящие. Использование примитивных «орудий» (палки, камни, кости). Дальнейшее развитие стадности |
| 25 млн. лет | Общие предки человекообразных обезьян и людей — дриопитеки | Древесный образ жизни. Стадность |
Кратко:
· Дриопитеки
· Австралопитеки
· Человек умелый (Homo Habillis)
· Человек прямоходящий (Homo erectus)
· Человек неандертальский (Homosapiensneanderthalensis)
· Человек разумный (Homosapienssapiens)
Соотношение биологического и социального в человеке на разных этапах его становления:
· Дриопитеки
o Мозг 450 – 500 куб. см
o были представлены несколькими видами
o выраженный половой диморфизм
o Использовали предметы, но не изготовляли орудий труда
· Человек умелый (Homo Habillis)
o Мозг 650 куб. см
o Основной признак - «чопперы» - примитивные орудия из гальки – галечная культура
o Возможно, строили жилища
o На стоянках множество расколотых костей животных –наши предки доедали добычу за хищниками, раскалывая кости и доставая мозг
· Человек прямоходящий (Homo erectus)
o Мозг 1000 куб. см
o Вовсю охотились, но ели и др. др.
o Могли использовать огонь
o Имели примитивную речь
· Человек неандертальский (Homosapiensneanderthalensis)
o Мозг как у нас – 1300 – 1400 куб. см, но форма другая – низкий свод черепа, надглазничный валик, шишковидный затылок
o Сложные формы охоты
o Огонь, жилища и одежда
o Захоронения
o Зачатки религии
· Человек разумный (Homosapienssapiens)
o Мозг 1300 – 1400
o Заселил все биотопы Земного шара
o Сельское хозяйство
Методы изучения эволюции человека (выделено жирным):
Изучение 1-го этапа производят методами палеонтологии и сравнительной анатомии. В связи с появлением элементов материальной культуры 2-й этап изучают также методами археологии. На 3-м этапе эволюционные события происходят в основном на молекулярно-генетическом уровне и проявляются на популяционном уровне. Основными способами изучения эволюции современного человека, поэтому являются биохимический, цитогенетический, популяционно-статистический методы и т.д. Изучение первых двух этапов эволюции человека сталкивается с необходимостью датировки палеонтологического материала и элементов материальной культуры.
Для определения абсолютного возраста ископаемых остатков человека и его предков широко используют физические методы, в частности радиометрические.С помощью масс-спектрометров определяют изотопный состав изучаемого объекта и по соотношению элементов с учетом периода полураспада входящих в его состав радиоактивных изотопов выявляют возраст образца.
Ископаемый костный материал содержит в своем составе минеральные компоненты и белок коллаген, разрушающийся чрезвычайно медленно. На этом основан коллагеновый метод абсолютной датировки ископаемых остатков чем меньше коллагена содержится в образце, тем более велика его древность. В последнее время в антропологии все более активно применяют методы иммунологии, молекулярной биологии и цитогенетики. В связи с огромной важностью этих методов остановимся на них подробнее.
Для определения прямого родства организмов друг с другом используют иммунологический метод, основанный на изучении иммунологических реакций антиген - антитело.Его можно применять для изучения степени родства не только современного человека с человекообразными обезьянами, но и ныне живущих видов с ископаемыми. Для этого следовые количества белка, извлекаемые из костей ископаемых форм, используют для получения антител, которые и применяют в иммунных реакциях с белками современных видов.
85. Характеристика основных этапов антропогенеза. Соотношение биологического и социального в человеке на разных этапах его становления.
Биологическая эволюция – биологическая форма движения материи;
· Как биологический вид человек относится к отряду Приматы;
Социальная эволюция – эволюция общества;
Этапы:
1) Досапиентный этап – историческое развитие эволюционных предков современных людей;
- В соответствии с закономерностями и эмпирическими правилами биологической эволюции;
Ø Закономерности: полифункциональность органов/структур, изменение количества отдельных функций, расширение и смена функций;
Ø Эмпирические правила: происхождение от наименее специализированных предковых форм, необратимость процесса;
- Примерно 22 млн лет назад и позже, появились признаки бипедии и зачатки предметно-орудийной деятельности;
2) Этап Сапиентации – обособление рода Homo, завершающая стадия исторического развития эволюционных предков современных людей и становление современного человечества;
- Соответственно закономерностям Социальной эволюции (включает Социальные революции);
Пусковой механизм Антропогенеза – «первая культурная революция», итог которой – превращение культуры из маловажного и необязательного дополнения к естественным видам поведения животных в адаптивный фактор, определяющий и организующий главные стороны жизнедеятельности гоминид;
На Досапиентном этапе выделялись линии эволюции, давшие современных человекообразных обезьян в примерной последовательности: Гиббон, орангутан, горилла, шимпанзе;
Причины выделения человеческой линии эволюции:
- Эколого-адаптационная гипотеза: изменение среды жизни и фундаментальные сдвиги социального поведения в условиях похолодания в верхнем миоцене (45-25 млн лет назад);
- Генетическая гипотеза: мутация регуляторных генов и хромосомные перестройки, ставшие причиной изменения онтогенеза (набор структурных генов у людей и шимпанзе практически идентичен, 24 пары хромосом шимпанзе и 23 человека, при этом хромосома 2 человека точно соответствует двум соединенным хромосомам шимпанзе);
86. Внутривидовая дифференциация человечества. Расы. Концепции классификации и происхождения рас. Расы и генетический полиморфизм человека.
С момента возникновения Н. sapiens социальное в человеке стало его сущностью и биологическая эволюция видоизменялась, проявляясь в возникновении широкого генетического полиморфизма.
Генетическое разнообразие на уровне генов и в меньшей степени хромосом обеспечивает разнообразие генотипов особей. Разнообразные генотипы по-разному проявляются в меняющихся условиях среды, давая огромное фенотипическое многообразие людей.
В основе морфофизиологического полиморфизма человечества лежат полиморфизм наследственного материала на уровне генома и модификационная изменчивость. Эти факторы обеспечивают не только индивидуальное морфофизиологическое многообразие, но и внутривидовую групповую дифференциацию человечества на расы и адаптивные экологические типы.
Раса — биологические подразделения человечества современного вида (Homo sapiens), различающиеся общими наследственными морфологическими особенностями, связанными с единством происхождения и определенной областью обитания.
Одним из первых создателей расовой классификации был французский ученый Франсуа Бернье, опубликовавший в 1684 г. работу, в которой употребил термин «раса». Антропологи выделяют четыре большие расы первого порядка и ряд промежуточных, численно небольших, но также самостоятельных. Кроме того, в каждой расе первого порядка выделяются основные подразделения —
Негроидная раса: негры, негрилли, бушмены и готтентоты.
Характерные черты негроида:
- курчавые волосы (черные);
- темно-коричневая кожа;
- карие глаза;
- слабое развитие третичного волосяного покрова;
- умеренно выступающие скулы;
- сильно выдающиеся челюсти;
- толстые губы;
- широкий нос.
Смешанные и переходные формы между негроидной и европеоидной большими расами: эфиопская раса, переходные группы Западного Судами, мулаты, «цветные» африканские группы.
Европеоидная раса: северные, переходные формы, южные.
Характерные черты европеоида:
- волнистые или прямые мягкие волосы разных оттенков;
- светлая или смуглая кожа;
- карие, светло-серые и голубые глаза;
- слабовыступающие скулы и челюсти;
- узкий нос с высоким переносьем;
- тонкая или средняя толщина губ. Смешанные формы между европеоидной большой расой и американской ветвью монголоидной большой расы: американские метисы.
Смешанные формы между европеоидной большой расой и азиатской ветвью монголоидной большой расы: среднеазиатские группы, южносибирская раса, лапоноиды и субуральский
Монголоидная раса: американские расы, азиатская ветвь монголоидных рас, континентальные монголоиды, арктическая раса (эскимосы и палеоазиаты), тихоокеанские (восточноазиатские) расы.
Характерные черты монголоида:
- прямые, жесткие и темные волосы;
- слабое развитие третичного волосяного покрова;
- желтоватый оттенок кожи;
- карие глаза;
- уплощенное лицо с сильно выделяющимися скулами;
- узкий нос, часто с низким переносьем;
- наличие эпикантуса (складка у внутреннего утла глаза).
Переходные группы между азиатской ветвью монголоидной большой расы и австрало- идной большой расой: южноазиатская раса (южные монголоиды), японцы, восточноиндонезийская Рис. 3.3. Монголоидная группа
Австралоидная раса: веддоиды, австралийцы, айны, папуасы и меланезийцы, негритосы. Характерные черты австралоида:
- темная окраска кожи;
- карие глаза;
- широкий нос;
- толстые губы;
- волнистые волосы;
- сильцо развит третичный волосяной покров.
Другие расовые типы (смешанные): малагасийцы, полинезийцы, микронезийцы, гавайцы.
Существуют две основные гипотезы происхождения человеческих рас — полицентризма и моноцентризма.
Согласно теории полицентризма, современные расы человека возникли в результате длительной параллельной эволюции нескольких филетических линий на разных материках: европеоидная в Европе, негроидная в Африке, монголоидная в Центральной и Восточной Азии, австралоидная в Австралии. Однако если эволюция расовых комплексов и шла параллельно на разных континентах, она не могла быть полностью независимой, поскольку древние проторасы должны были скрещиваться на границах своих ареалов и обмениваться генетической информацией. В ряде областей сформировались промежуточные малые расы, характеризующиеся смешением признаков разных больших рас уже в древности.
С позиций моноцентризма современные человеческие расы сформировались относительно поздно, 30-35 тыс. лет назад, в процессе расселения неоантропов из области их возникновения. При этом также допускается возможность скрещивания (хотя бы ограниченного) неоантропов во время их экспансии с вытесняемыми популяциями палеоантропов (как процесса интрогрессивной межвидовой гибридизации) с проникновением аллелей последних в генофонды популяций неоантропов. Это также могло способствовать расовой дифференциации и устойчивости некоторых фенотипических признаков. Существуют и компромиссные между моно- и полицентризмом концепции, допускающие расхождение филетических линий, ведущих к разным большим расам, на различных уровнях (стадиях) антропогенеза: например, более близких друг к другу европеоидов и негроидов уже на стадии неоантропов с первоначальным развитием их предкового ствола в западной части Старого Света, тогда как еще на стадии палеоантропов могла обособиться восточная ветвь — монголоиды и, может быть, австралоиды, хотя по некоторым отдельным признакам европеоиды имеют общие признаки и с австролоидами.