
Нейроны ЦНС: классификация, их функциональные структуры, величина потенциала покоя и потенциала действия. Классификация синапсов ЦНС, физиологическая оценка синапсов разных классов, медиаторы синапсов ЦНС и их функциональное значение.
Классификация нейронов.
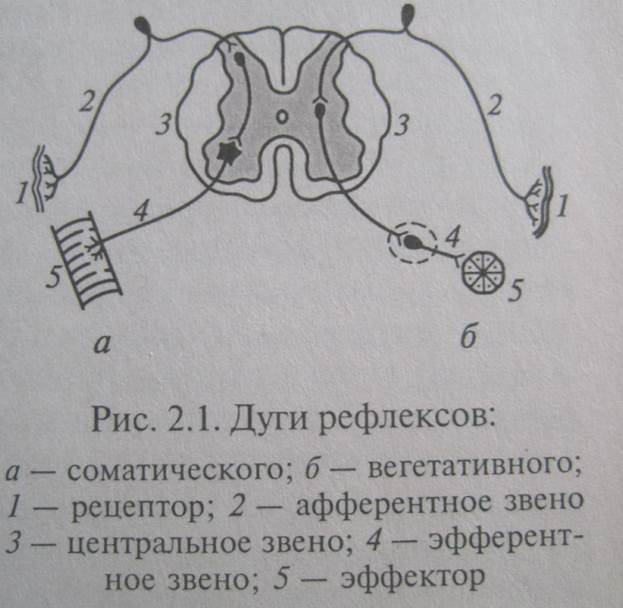
По обеспечению соматических или висцеральных функций подразделяются на соматические и вегетативные соответственно двум отделам нервной системы. По месту в системе рефлекторных связей (соответственно по направлению или источнику информации) на афферентные, воспринимающие с помощью сенсорных рецепторов инфу о внешней и внутренней среде организма и передающие ее в вышележащие отделы ЦНС; эфферентные, передающие инфу к рабочим органам – эффекторам; вставочные, обеспечивающие взаимодействие между нейронами ЦНС. По хим природе медиатора, выделяющегося в окончаниях аксонов, адренергические, холинергические, серотонинергические и др. По типу ответной реакции эффектора тормозящие и возбуждающие. Нейроны высших отделов ЦНС по специфичности воспринимаемой сенсорной информации могут быть моно, би и полисенсорными. По характеру активности фоноактивные и молчащие, возбуждающимися только в ответ на раздражение. Фоноактивные нейроны играют большую роль в поддержании тонуса ЦНС и особенно коры большого мозга.
Классификация синапсов. В зависимости от местоположения аксо – соматические, аксодендритные, аксоаксонные, сома – соматические, дендросоматические, дендродендритные. По рабочему эффекту возбуждающие и тормозящие. По способу передачи сигналов химические, электрические, смешанные.
Свойства химических синапсов.
Большинство синапсов являются химическими. Первое их преимущество заключается в том, что хим синапсы благодаря наличию различных медиаторов и постсинаптических мембран могут оказывать возбуждающие и тормозное влияние на нервные и эффекторные клетки. Второе – могут передавать инфу в широких временных рамках. Третье – высокая пластичность; их функция зависит от предшествующей нейрональной активности, поэтому они играют важную роль в процессах обучения и памяти.
Химические синапсы. На пресинаптической поверхности активной зоны сос торны синаптической везикулы находятся филаменты и частицы, выступающие из пресинаптической терминали, часто формирующие правильный шестиугольник, названный пресинаптической решеткой. Определенные точки на сетке являются участками экзоцитоза синаптических везикул.
Медиатор в пресинаптическом окончании находится в двух пулах – готовом к экзоцитозу (пул 1 – медиатор лежит на пресинаптической мембране изнутри) и мобилизационном (пул 2 – находится в цитоскелете), из которого пополняется в течение нескольких секунд пул 1. Этоо процесс активируется ионами кальция. Химическим синапсам свойственно явление суммации.
Электросекреторное сопряжение в ЦНС обеспечивается ионами кальция: при возбуждении пресинаптической терминали открываются потенциалзависимые кальциевые каналы. Кальций входит в нервное окончание согласно электрохимическому градиенту и взаимодействует с экзоцитозным аппаратом пресинапса, представляющим собой совокупность белков.
Активация белков кальцием вызывает выброс медиатора в синаптическую щель. Синаптическая щель хим синапсов содержит поперечно расположенные гликопротеиновые филаменты, обеспечивающие связи пре и постсинаптической мембран и направление диффундирующего медиатора. Медиатор взаимодействует с белком – рецептором постсинаптической мембраны, и эффекторная клетка приходит в состояние возбуждения или торможения.
Электрические возбуждающие синапсы. Обнаружены в составе мезенцефального ядра тройничного нерва, вестибулярного ядра Дейтерса, ядра нижней оливы продолговатого мозга. Характерны для сердца, гладких мышц, секреторных клеток. Имеются следующие электрические синапсы: аксон – сома, аксон – дендрит, аксон – аксон, дендрит – дендрит, дендрит – сома, сома – сома.
Проводят сигнал в обе стороны без синаптической задержки, передача не блокируется при удалении кальция. Контактирующие мембраны клеток в составе электрического синапса связаны друг с другом полуканалами белковой природы – коннексонами. Через коннексоны клетки обмениваются аминокислотами, пептидами, РНК, метаболитами, циклическими нуклеотидами.
Механизм передачи возбуждения в электрическом синапсе: ПД, возникающий на мембране одной клетки, с помощью своего электрического поля деполяризует мембрану соседней клетки до КП.
Механизм возбуждения нейронов ЦНС: реакция нейрона на одиночный импульс и серию импульсов, возбуждающий постсинаптический потенциал (ВПСП) и его свойства, роль кальция, место возникновения ВПСП и потенциала действия, роль дендритов.
Механизм возбуждения нейронов.
1. В возникновении ПД нейронов принимают участие ионы кальция, ток которых в клетку более медленный, чем ток натрия (натрий вносит основной вклад в возникновение ПД).
2. Для возникновения ПД в нейроне необходим поток афферентных импульсов. Один ПД, пришедший в пресинаптическое окончание, обеспечивает выделение 1 – 20 квантов медиатора. При этом возникает небольшой ВПСП. Выброс медиатора обеспечивает входящий в дерполяризационную терминаль кальций. При поступлении импульсов к постсинаптическому нейрону в результате суммации ВПСП возникает ПД постсинаптического нейрона. ВПСП возникает вследствие суммарного тока ионов в клетку и из клетки через ионные каналы по электрохим градиенту.
Поступившей в пресинаптическое окончание кальций удаляется из него при помощи кальциевого насоса. Прекращение действия медиатора осуществляется посредством обратного захвата пресинаптическим аппаратом, с помощью разрушения специальными ферментами. Норадреналин разрушается монаминоксидазой и катехолметилтрансферазой, АХ гидролизуется ацетилхолинтрансферазой. Прекращение действия избытка медиатора на постсинаптическую мембрану предотвращает десенситизацию – снижение чувствительности постсинаптичекой мембраны к действующему медиатору.
Пептидные медиаторы, ферменты и другие белки транспортируются в пресинаптические окончания из тела клетки по аксону с помощью микротрубочек и микрофиламентов, тянущихся по всей длине аксона. Для этого транспорта необходимы кальций и энергия.
3. Место возникновения генераторных ВПСП - тело нейрона. Поскольку постсинаптические мембраны синапсов находятся в непосредственной близости от места возникновения ПД, располагающегося на аксонном холмике. Близость к аксонному холмику ВПСП, возникающих в теле нейрона, обеспечивает участие их в генерации ПД.
4. Место возникновения ПД нейрона: формирование ПД начинается на мембране аксонного холмика. Синапсы на нем отсутствуют, возбудимость мембраны в 3 – 4 раза выше возбудимости мембраны тела нейрона, что объясняется более высокой концентрацией натриевых каналов. ВПСП, возникающие в любых участках тела нейрона суммируются, их суммарное электрическое поле может деполяризовать мембрану на расстоянии 1 – 2 мм. Электрическое поле начального возбуждения аксонного холмика обеспечивает деполяризацию тела нейрона до КП в результате чего возникает ПД нейрона, который проводится по аксону к др клетке. На восходящей части ПД нейрона регистрируется перегиб на уровне 30 - 40 мВ (задержка в развитии ПД) вследствие того, что возбудимость мембраны тела нейрона в 3 – 4 раза ниже возбудимости мембраны аксонного холмика. ПД нейрона – единый одновременный процесс возбуждения аксонного холмика и тела нейрона.
5. Роль дендритов в возникновении возбуждения нейрона: дендритные синапсы называются модуляторными. Они удалены на значительное расстояние от генераторного пункта нейрона – аксонного холмика. Суммарное электрическое поле их ВПСП не может вызвать достаточную деполяризацию аксонного холмика и обеспечить возникновение ПД, а определяют лишь возбудимость нейрона.