ФИЗИОЛОГИЯ НЕРВОВ, СИНАПСОВ, МЫШЦ И РЕЦЕПТОРОВ
Понятия возбудимость и раздражимость, возбудимые и невозбудимые ткани. Раздражители: определение, их виды, характеристика. Открытие животного электричества. Мембранный потенциал покоя: его схема, параметры, механизм формирования.
Возбудимость – способность клетки генерировать ПД при ее раздражении.
Раздражимость – общее свойство живой материи активно изменять свое состояние при действии раздражителя.
Возбудимые ткани – мышечная и нервная.
Невозбудимые – эпителиальная и соединительная.
Раздражитель – любое изменение внешней или внутренней среды организма, воспринимаемой клетками и вызывающее ответную реакцию.
Выделяют физические (электрические, механические, термические, световые) и химические. Также подразделяют на адекватные и неадекватные.
Начало исследованиям биоэлектрических потенциалов положил в конце 18 века Луиджи Гальвани. В первом опыте, подвешивая препарат обнаженных задних лапок лягушек с помощью медного крючка на железной решетке, он обнаружил, что при касании решетки мышцы сокращались. Ученый высказал предположение, что сокращение мышц является следствием воздействия на них электричества, источником которого являются животные ткани – мышцы и нервы. Однако Вольта оспорил данное утверждение. По его мнению, причиной сокращения был электрический ток, возникающий в области контакта двух разнородных металлов с тканями лягушки. Гальвани поставил второй опыт, в котором нерв нервно – мышечного препарата набрасывался на мышцу стеклянным крючком так, чтобы он касался поврежденного и неповрежденного участка мышцы. В этом случае мышца также сокращалась. Второй опыт считается абсолютным доказательством существования животного электричества.
ПП – относительно стабильная разность электрических потенциалов между наружной и внутренней средой клетки. Величина от -30 до -90 мВ (в скелетных мышцах до -90, у нейронов -60, у гладких мышц -30…-70, в сердечной мышце -80…-90).
Механизм формирования ПП.
Причиной формирования ПП является разная концентрация анионов и катионов внутри и вне клетки.

Потенциал действия: понятие, схема потенциала действия (фазы), его параметры, механизм возникновения. Восстановительный период.
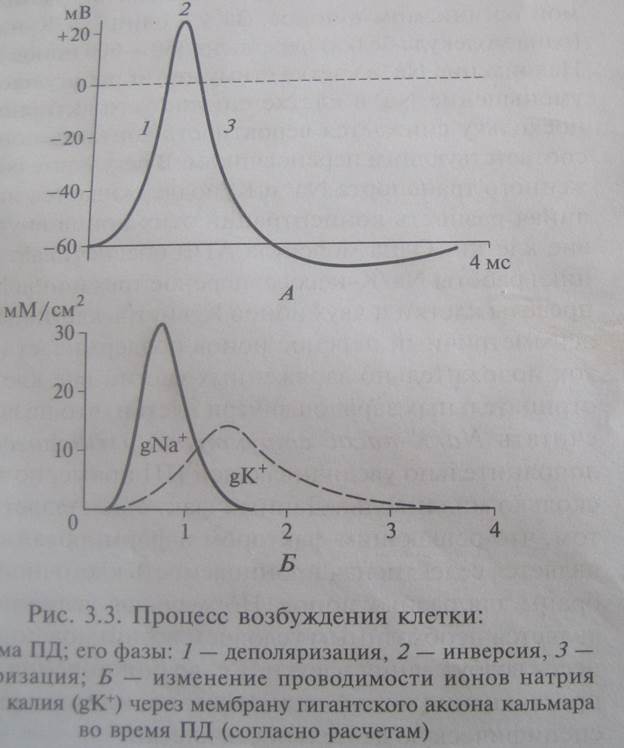
ПД – быстрое колебание величины мембранного потенциала вследствие активации и инактивации ионных каналов и диффузии ионов в клетку и из клетки. Величина колеблется от 80 до 130 мВ (у нервного волокна 110, у мышечного 120 – 130).
ПД состоит из двух основных фаз: деполяризации и реполяризации.
Механизмы
Ионные каналы
Если в формировании потенциала покоя участвуют постоянно открытые калиевые каналы без ворот, то в формировании ПД — два вида потенциалчувствительных каналов:
¾быстрые натриевые каналы;
¾потенциалчувствительные калиевые каналы.
Быстрые натриевые каналы абсолютно необходимы для возникновения ПД и определяют все его основные свойства (закон «все или ничего», распространение без затухания, наличие рефрактерности; см. выше, разд. «Физиологический смысл биопотенциалов»); потенциалчувствительные калиевые каналы играют вспомогательную роль.
Быстрые натриевые каналы
Они обладают двумя воротами:
¾наружными — активационными;
¾внутренними — инактивационными.
Будучи потенциалчувствительными, быстрые натриевые каналы обладают потенциалзависимостью и времязависимостью (рис. 1.8, А):
¾потенциалзависимость: активационные ворота при потенциале покоя закрыты, инактивационные — открыты; в ответ на деполяризацию активационные ворота открываются, инактивационные — закрываются;
¾времязависимость: активационные ворота в ответ на деполяризацию открываются чрезвычайно быстро (за десятые доли миллисекунды), инактивационные закрываются также достаточно быстро, но все же медленнее.
Таким образом,
¾в состоянии покоя быстрые натриевые каналы закрыты (так как закрыты активационные ворота);
¾в ответ на деполяризацию быстрые натриевые каналы сначала быстро открываются (инактивационные ворота еще не закрылись, активационные уже открылись); это так называемая натриевая активация;
¾затем с некоторой задержкой эти каналы закрываются (так как в ответ на деполяризацию закрываются инактивационные ворота); это так называемая натриевая инактивация.
Потенциалчувствительные калиевые каналы
Они обладают только наружными (активационными) воротами.
Данные каналы также характеризуются потенциалзависимостью и времязависимостью (рис. 1.8, Б):
¾потенциалзависимость: ворота при потенциале покоя закрыты, а в ответ на деполяризацию открываются;
¾времязависимость: ворота в ответ на деполяризацию открываются сравнительно медленно (за миллисекунды).
Таким образом,
¾в состоянии покоя потенциалчувствительные калиевые каналы закрыты;
¾при деполяризации потенциалчувствительные калиевые каналы сравнительно медленно открываются (калиевая активация);
¾при продолжающейся деполяризации эти каналы остаются открытыми и закрываются лишь тогда, когда мембранный потенциал возвращается к уровню покоя.
Фаза деполяризации
Фаза деполяризации ПД обусловлена входом в клетку Na+. Действительно, деполяризация может быть обусловлена только входом катиона,а из двух катионов, участвующих в создании биопотенциалов (Na+ и K+), только Na+ стремится войти в клетку. Из этого следует, что в фазу деполяризации натриевая проницаемость должна резко повысится — то есть должны открыться быстрые натриевые каналы. Эти каналы, как уже говорилось, открываются в ответ на деполяризацию. Следовательно, деполяризация является единственным раздражителем, способным вызвать ПД.
Итак, с одной стороны, деполяризация вызывает открывание натриевых каналов, а с другой — вход Na+ по этим каналам вызывает деполяризацию. Таким образом, вход Na+ в фазу деполяризации ПД носит самоусиливающийся характер: деполяризация приводит к открыванию натриевых каналов; в результате входит Na+; вход Na+ приводит к еще большей деполяризации; нарастающая деполяризация вызывает открывание новых натриевых каналов и т.д.
Фаза реполяризации
На пике ПД движущие силы, проницаемости и потоки Na+ и K+ меняются следующим образом.
· Для Na+:
¾движущая сила (электрохимический градиент) резко падает, так как клетка становится заряженной положительно изнутри, и электрический градиент для Na+ становится направленным наружу (противоположно концентрационному);
¾проницаемость также падает, так как закрываются инактивационные ворота быстрых натриевых каналов;
¾следовательно, входящий ток Na+ резко снижается.
· Для K+:
¾движущая сила (электрохимический градиент) резко возрастает, так как клетка становится заряженной положительно изнутри, и электрический градиент для K+ становится направленным наружу (как и концентрационный);
¾проницаемость, будучи большой уже в покое, дополнительно возрастает, так как открываются потенциалчувствительные калиевые каналы;
¾следовательно, выходящий ток K+ резко повышается.
Таким образом, фаза реполяризации ПД обусловлена выходом K+ на фоне прекращающегося входа Na+.
Следовые потенциалы
По окончании ПД часто возникают так называемые следовые потенциалы, в большинстве случаев обусловленные тем, что ворота ионных каналов не успевают вернуться в состояние, характерное для потенциала покоя. Если следовой потенциал положительнее потенциала покоя, то он называется следовой деполяризацией, если отрицательнее — следовой гиперполяризацией. Эти потенциалы в разных клетках разные; в некоторых клетках их нет вовсе, в других может быть несколько сменяющих друг друга следовых потенциалов. В качестве примера на рис. 1.10 приведена следовая гиперполяризация.Видно, что она обусловлена временным повышением калиевой проницаемости (из-за медленного закрывания потенциалчувствительных калиевых каналов) и, как следствие, смещением мембранного потенциала в сторону EK.
Фазовые изменения возбудимости ткани во время ее возбуждения - потенциала действия (график), их механизм. Сравнительная характеристика локального потенциала и потенциала действия. Закон “все или ничего“.
1. Во время локального ответа возбудимость повышена. Причина: мембранный потенциал приближается к Eкр.
2. Во время фазы деполяризации, пика ПД и первой трети фазы реполяризации возбудимость падает до нуля (никакие раздражители не способны вызвать повторный ПД). Это период абсолютной невозбудимости, или абсолютной рефрактерности — одно из ключевых свойств ПД (см. выше, разд. «Физиологический смысл биопотенциалов»). Причина: закрываются инактивационные ворота быстрых натриевых каналов, и в этих условиях никакие раздражители не способны вызвать открывание этих каналов.
3. Начиная от второй трети фазы реполяризации возбудимость постепенно восстанавливается, хотя и остается сниженной по сравнению с исходной (ПД может быть вызван, но только сильными раздражителями). Это период относительной невозбудимости, или относительной рефрактерности. Причина: постепенно вновь открываются инактивационные ворота быстрых натриевых каналов. Если следовых потенциалов нет, то возбудимость восстанавливается почти одновременно с окончанием фазы реполяризации (рис 1.14, А). Если же имеются следовые потенциалы, то изменения возбудимости более сложны. В частности, во время следовой гиперполяризациивозбудимость понижена, поэтому период относительной рефрактерности длится дольше — до окончания следовой гиперполяризации (рис. 1.14, Б). Причины: 1) мембранный потенциал смещен в сторону более отрицательных значений; 2) калиевая проницаемость повышена, и поэтому Eкр смещен в сторону более положительных значений (рис. 1.13, В). Обе эти причины приводят к росту порога деполяризации, и, следовательно, снижению возбудимости.
Каковы бы ни были особенности изменений возбудимости в отдельных нейронах, главной и важнейшей особенностью этих изменений является наличие абсолютной рефрактерности: почти все время, пока длится ПД, повторный ПД вызвать невозможно.
От длительности периода абсолютной рефрактерности зависит, сколько ПД в 1 секунду может возникнуть в той или иной клетке. Например, если рефрактерность составляет 1 мс, то максимальная частота ПД равна около 1000 в 1 с, то есть 1000 Гц. Эта максимальная частота импульсов, которую может воспроизвести та или иная ткань, клетка или часть клетки, является мерой лабильности («функциональной подвижности ткани»).


Критерии для оценки возбудимости ткани. Явление аккомодации возбудимой ткани. Лабильность: понятие, мера лабильности, факторы ее определяющие, схема эксперимента для определения лабильности нерва, синапса и мышцы.
Возбудимость клетки оценивается несколькими параметрами.
1. Пороговый потенциал – минимальная величина, на которую надо уменьшить МПП, чтобы вызвать возбуждение в виде ПД. Небольшая величина порогового потенциала свидетельствует о высокой возбудимости клетки.
2. Пороговая сила – наименьшая сила раздражителя, способная вызвать возбуждение при неограничении времени ее действия. Реобаза – наименьшая сила электрического тока, способная вызвать импульсное возбуждение при неограничении времени раздражения. Большая пороговая сила свидетельствует о низкой возбудимости.
3. Пороговое время – минимальное время, в течение которого должен действовать на ткань раздражитель пороговой силы, чтобы вызвать ее возбуждение. Хронаксия – наименьшее время, в течение которого должен действовать электрический ток в две реобазы, чтобы вызвать возбуждение.
Аккомодация — это снижение возбудимости при уменьшении крутизны (скорости нарастания) раздражителя. Причина аккомодации — та же, что и длительной деполяризации: при медленно нарастающей деполяризации успевают развиться натриевая инактивация и калиевая активация.
Лабильность – функциональная подвижность, скорость протекания отдельных циклов возбуждения в возбудимых тканях.
Мера лабильности – максимальное количество импульсов, которое может воспроизвести ткань в единицу времени в соответствии с ритмом возбуждения.
Лабильность определяется скоростью перемещения ионов в клетку и обратно, и она в свою очередь зависит от скорости изменения проницаемости клеточной мембраны. Чем больше рефрактерная фаза, тем ниже лабильность ткани.
Нервное волокно: функциональное значение отдельных структурных элементов, классификация нервных волокон, механизм проведения возбуждения по мякотным и безмякотным волокнам, закономерности проведения возбуждения по нервному волокну и опыты их доказывающие.
Нервные волокна – отростки нейронов, с помощью которых осуществляется связь нейронов между собой, а также с иннервируемыми клетками и рецепторами.
Оболочку безмиелиновых волокон образуют швановские клетки, в которые погружаются осевые цилиндры нейронов. Оболочку миелиновых волокон образуют в периферической НС леммоциты, а в ЦНС олигодендроциты. Миелиновая оболочка влияет на электрические свойства волокна, способствует увеличению скорости проведения возбуждения, выполняет трофическую функцию (регуляция метаболизма, рост осевого цилиндра)
Плазматическая мембрана определяет пассивные электрические свойства волокна (сопротивление и емкость мембраны). Распределение и концентрация электроуправляемых каналов влияет на возбудимость и проводимость нервного волокна.
Нейрофибрилы и микротрубочки обеспечивают аксональный транспорт.
Появление разности потенциалов между возбужденными и невозбужденными участками мембраны. =>возникновение локальных токов между возбужденными и невозбужденными участками мембраны=>генерация новых ПД в соседних участках волокна.
Законы проведения возбуждения по нервному волокну
1. Анатомической и физиологической целостности
2. Изолированное проведение возбуждения
3. Двустороннее проведения возбуждения
Если в эксперименте нанести раздражение в любой участок нервного волокна, то возбуждение регистрируется как в проксимальном, так и в дистальном участке нерва
4. Относительная неутомляемость нервного волокна
Нервно-мышечный синапс: доказательства его наличия, назначение отдельных структурных элементов, механизм проведения возбуждения, особенности проведения в синапсе по сравнению с нервным волокном и опыты, их доказывающие.
Синапс – специализированная структура, обеспечивающая передачу сигнала от клетки к клетке с помощью химических веществ или электрического поля.
В состав синапса входит пре и постсинаптическая мембраны и синаптическая щель.
В области пресинаптической мембраны находятся многочисленные везикулы, которые содержат молекулы медиатора ацетилхолина. Везикулы концентрируются в особых областях- активных зонах.
Постсинаптическая мембрана образует постсинаптические складки, на гребнях которых располагаются АХ – рецепторы.
Синаптическая щель заполнена межклеточной жидкостью, в ней располагается базальная пластинка, которая содержит различные белки. Они способствуют плотному сцеплению пре и постсинаптических мембран. Базальная мембрана содержит фермент ацетилхолинэстераза, котрая разрушает высвобожденный ацетилхолин и готовит синапс к проведению очередного сигнала.
Механизм синаптической передачи.
1. Выброс медиатора в синаптическую щель. Он запускается посредством ПД пресинаптического окончания. Деполяризация мембраны пресинаптического окончания ведет к открытию потенциал управляемых Ca – каналов. Ca входит в нервное окончание согласно электрохимическому градиенту. Са активирует экзоцитозный аппарат пресинапса, что обеспечивает выброс ацетилхолина посредством экзоцитоза в синаптическую щель
2. Диффузия АХ к постсинаптической мембране, где локализуются N – холинорецепторы.
3. Возникновение возбуждения в мышечном волокне. В результате взаимодействия АХ с N – холинорецепторами открываются ионные каналы и вследствие преобладания входа Na в клетку происходит деполяризация постсинаптической мембраны.
4. Удаление АХ из синаптической щели. Осуществляется путем разрушения под действием АХЭ, около 60 % холина захватывается обратно пресинаптическим окончанием, часть АХ рассеивается.
Особенности проведения в синапсе по сравнению с нервным волокном.
1. Задержка в передаче сигнала к другой клетке. Это время затрачивается на высвобождение медиатора из нервного окончания, диффузию его к постсинаптической мембране и возникновение постсинаптических потенциалов, способных вызвать ПД в мышечном волокне.
2. Низкая лабильность синапса. Она объясняется тем, что необходимо время для высвобождения медиатора, распространения его до постсинаптической мембраны, возникновение ВПСП, а при более частом раздражении в синапсе возникают длительные ВПСП. Суммируясь они могут вызвать стойкую деполяризацию постсинаптичекой мембраны, что делает невозможным генерацию новых ПД, т.к. ПД может возникнуть только при наличии заряда в клетке.
3. Одностороннее проведение. Возбуждение передается только от нервного волокна к другой клетке, т.к. пресинаптическое окончание чувствительно только к нервному импульсу, а постсинаптическая мембрана к медиатору.
4. Неизолированное проведение. Возбуждение рядом расположенных постсинаптических мембран суммируется
5. Затухание возбуждения в химических синапсах принедостаточном выделении медиатора из пресинаптического окончания в синаптическую щель
6. Проводимость нервно – мышечного синапса угнетается и стимулируется различными веществами.
7. Утомляемость синапса. Главная причина – истощение запасов медиатора в пресинаптическом окончании.
7. Скелетная мышца: значение отдельных структурных элементов мышечного волокна, понятие о структурной и функциональной единице изолированной мышцы и двигательного аппарата организма, классификация двигательных единиц, физиологические свойства скелетной мышцы и ее функции
Структурной и функциональной единицей мышцы, является мышечное
волокно, представляющее собой сильно вытянутое многоядерное волокно.
Мышечное волокно имеет оболочку сарколемму. В саркоплазме мышечного
волокна имеются многочисленные ядра, митохондрии, растворимые белки,
капельки жира, гранулы гликогена, фосфатсодержащие вещества, другие
малые молекулы, а также ионы. Там же расположены многочисленные
мембраны. Они включены в состав поперечных Т – трубочек, пересекающих
мышечное волокнои связывающих сарколемму с внутриклеточным
пространством и саркоплазматическим ретикулумом. Последний
представляет собой систему связанных друг с другом систем и канальцев. Т –
система и ретикулум обеспечивают функциональное согласование процессов
возбуждения клеточной мембраны со специфической активностью
сократительного аппарата миофибриллы. Через Т – трубочки может
происходить выделение продуктов обмена в межклеточное пространство и
далее в кровь.
Внутри мышечного волокна тянется масса миофибрилл, с которыми связана
способность мышц к сокращению.
Двигательная единица – комплекс мотонейрона со всеми его коллатералями и группой иннервируемых им мышечных волокон.
Двигательные единицы подразделяют на быстрые и медленные. Медленные мотонейроны низкопороговые, малоутомляемые. Быстрые мотонейроны высокопороговые, утомляемые, не способны к длительному поддержания высокочастотного разряда.
Физиологические свойства скелетной мышцы.
1.Возбудимость. Способность мышцы отвечать на действие раздражителя самой мышцы или двигательного нерва изменением физиологических свойств и возникновением возбуждения.
2.Проводимость. Способностъ проводить возбуждение, возникшее в каком-либо участке мышечного волокна, по всему волокну.
3.Рефрактерность. Временное снижение возбудимости мышцы, которое возникает в результате возбуждения.
4.Лабильность. Количество возбуждений за единицу времени, зависящее от уровня обменных процессов.
5.Сократимость. Способность изменять свою длину или напряжение при возбуждении. Это основная функция скелетном мышцы. В период относительного покоя скелетные мышцы полностью не расслаблены, а умеренно напряжены. Такое состояние называется мышечным тонусом и объясняется редкими импульсами от двигательных нейронов, которые попеременно возбуждают нейромоторные единицы. При изотоническом сокращении укорачивается мышечное волокно, а напряжение не изменяется; при изометрическом сокращении длина мышцы не изменяется, а напряжение возрастает.
При этом мышцы выполняют следующие функции:
1) обеспечивают определенную позу тела человека;
2) перемещают тело в пространстве;
3) перемещают отдельные части тела относительно друг друга;
4) являются источником тепла, выполняя терморегуляционную функцию.