Наиболее часто в настоящее время используют два способа ДНК-типирования патогенных микроорганизмов, в основе которых лежит метод ПЦР. В первом случае используют один или несколько коротких праймеров произвольной первичной структуры длиной в 6–15 нуклеотидов, которые из-за своих малых размеров обладают низкой специфичностью в отношении конкретных генетических локусов и способны гибридизоваться со многими сайтами геномной ДНК. Во втором случае применяют специфические олигонуклеотидные праймеры длиной в 20–27 нуклеотидов, последовательности которых фланкируют исследуемую последовательность нуклеотидов в бактериальном геноме.
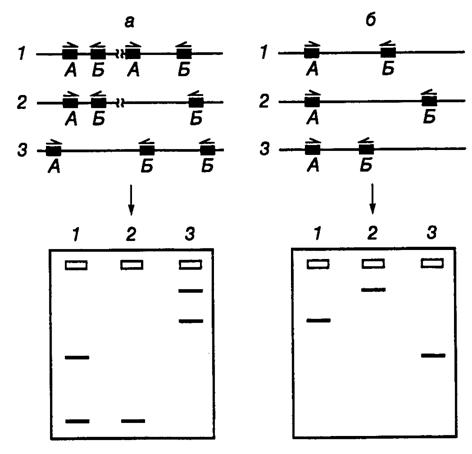
Рис. II.35. Схема ДНК-типирования микроорганизмов с использованием ПЦР и праймеров произвольной структуры
а – штаммоспецифические различия, обусловленные разной локализацией участков ДНК, взаимодействующих с праймерами. В ДНК штаммов 2 и 3 отсутствует участок, который имеется в ДНК штамма 1, что приводит к исчезновению соответствующей полосы продукта ПЦР на электрофореграмме; б – результаты амплификации ДНК, содержащей сайты связывания праймеров постоянной локализации. ДНК штаммов 2 и 3 содержат делеции разной длины между сайтами связывания праймеров. Альтернативно, ДНК штаммов 1 и 2 могут содержать вставки, отсутствующие у ДНК штамма 3. Это находит отражение в длинах продуктов ПЦР, разделяемых электрофорезом. А, Б – сайты связывания праймеров на ДНК
Генетическое типирование микроорганизмов с использованием праймеров произвольной первичной структуры. Использование коротких олигонуклеотидных праймеров произвольной структуры основано на том, что в больших геномах для них имеются множественные сайты посадки, а следовательно, и инициации ПЦР. Чем короче такие праймеры, тем большее количество сайтов посадки для них должно существовать. Одним из ограничений дальнейшего участия таких праймеров в ПЦР будет расстояние между двумя сайтами посадки для противоположно направленных праймеров. Чем длиннее ПЦР-продукт, который должен образовываться в результате функционирования таких праймеров, тем менее надежно работает вся система типирования. На практике длина образующихся с произвольными праймерами продуктов ПЦР находится в пределах 0,2–2,0 т.п.о.
В зависимости от локализации на ДНК мест посадки для пары коротких праймеров могут образовываться два типа продуктов ПЦР. В первом случае различия в длинах продуктов ПЦР обусловлены присутствием на ДНК типируемых микроорганизмов разного числа сайтов связывания для одного или обоих праймеров (рис. II.35, а). Во втором случае такие различия определяются длинами сегментов ДНК (ампликонов), заключенных между парами мест посадки праймеров при неизменном их числе в конкретных генетических локусах (см. рис. II.35, б). На практике могут одновременно реализовываться обе эти возможности. Во время генетического типирования эукариот указанными методами иногда сразу функционируют до 100 ампликонов, тогда как у бактерий это число достигает лишь 20.
Несмотря на то что при обсуждаемом подходе последовательности праймеров выбираются произвольно, получаемая картина амплификации, как правило, является видо- и штаммоспецифичной. При этом количество сайтов связывания на одной и той же ДНК для праймеров одинаковой длины, но с разной первичной структурой может существенно варьировать. На рис. II.35 показаны такие свойства двадцати 10-нуклеотидных праймеров, испытанных на ДНК человека, бобов и S. aureus. Видно, что использование праймера AGGGGTCTTG, например, приводит к амплификации 8 ампликонов в ДНК человека, 3 ампликонов в ДНК соевых бобов и не выявляет ни одного ампликона в ДНК S. aureus, тогда как при использовании праймера AATCGGGCTG удается обнаруживать до 40 ампликонов в ДНК человека и соевых бобов и до 20 ампликонов в бактериальной ДНК. Таким образом, при использовании обсуждаемого метода в ДНК-типировании самым важным этапом является подбор праймеров или их комбинаций для наиболее эффективного определения принадлежности организма к тому или иному виду, штамму или линии.
Генетическое типирование микроорганизмов с использованием геномных фингерпринтов. При другом подходе к генетическому типированию с использованием ПЦР исследуют полиморфизм конкретных локусов, для которых хотя бы частично известна первичная структура. Синтезируют специфические олигонуклеотидные праймеры, сайты связывания которых фланкируют исследуемую последовательность ДНК с длиной 1,5–2,5 т.п.о. После проведения ПЦР особенности первичной структуры продуктов ПЦР определяют с помощью рестрикционного анализа. Положение сайтов рестрикции в анализируемой последовательности нуклеотидов может иметь видо- или штаммоспецифический характер и служить точным генетическим маркером того или иного микроорганизма.
С помощью этого метода, который по своей сути является разновидностью рассмотренного выше способа определения ПДРФ, идентифицируют близкородственные, сходные по фенотипу штаммы возбудителей заболеваний, исследуют генетическую структуру популяций микроорганизмов и механизмы их адаптивной изменчивости.