Кроме поступающих из почвы воды и минеральных веществ, углеводов, образующихся в процессе фотосинтеза, необходимых в качестве источника энергии и строительных белков протоплазмы, растительная клетка для оптимального роста требует еще и некоторые другие химические соединения. К ним, в частности, относятся органические соединения – гормоны. Потребность в количестве гормонов обычно очень мала, и в большинстве случаев гормоны синтезируются в достаточных количествах самим растением.
Более того, между клетками в организме должен происходить обмен информацией. Одна группа клеток «отправляет» сигнал, другая – воспринимает его. Молекулу химической природы, обладающую сигнальной функцией, называют первичным мессенджером. Среди обширного спектра первичных мессенджеров выделяют и гормоны растений.
Признаки отнесения вещества к гормонам:
• вещество вызывает специфический физиологический ответ; особенность растительных гормонов в том, что они запускают крупные программы развития не только клеток, но и на уровне тканей, органов, целого растения;
• синтезируется в растении одной группой клеток, а отвечает за него другая группа (разобщено место синтеза и место действия, т.е. сигнальное вещество транспортируется). К синтезу гормонов потенциально способна любая клетка растений; как правило, фитогормоны являются низкомолекулярными соединениями;
• практически не играет роли в основном метаболизме клетки, а используется лишь для сигнальных целей.
• действует в низкой концентрации.
Первичные мессенджеры, воспринимаемые клеткой, специфически взаимодействуют со многими молекулами-мишенями, в частности с рецепторами. Чтобы клетка ответила на стимул, нужно включить внутриклеточную систему вторичных мессенджеров, усиливающих сигнал в десятки и сотни раз. Например, одна молекула ауксина активирует до 104 протонов.
Критерии обнаружения рецепторов:
• высокая избирательность и структурная специфичность в отношении гормона; чем больше физиологический ответ, тем выше аффинность[1];
• эффект проявляется при низких концентрациях гормона;
• кинетика связывания с агонистом описывается кривой с насыщением;
• при взаимодействии с рецептором не должна меняться химическая структура гормона;
• взаимодействие гормона с рецептором приводит к включению системы вторичных посредников.
Большинство систем вторичных мессенджеров, изученных у животных, обнаружены и у растений. Это фосфатидилинозитольная и аденилат- и гуанилатциклазные системы.
Рецепторы обычно локализованы в плазмалемме, но могут находиться и в других компартментах клетки. Рецептор связан с вторичными мессенджерами. Передача сигнала с помощью вторичных посредников активизирует процессы фосфорилирования/дефосфорилирования, что приводит к изменению метаболизма и работы цитоскелета.
Итак, любой гормон представляет собой вещество, образуемое в малых количествах в одной части организма и транспортируемое затем в другую часть растения, где он оказывает соответствующее действие. Расстояние, на которое транспортируется гормон, может быть относительно большим, например, от корня до листа, от последнего к почке, может быть и меньшим – от апикальной меристемы до находящихся ниже клеток, или совсем малым – в пределах одной клетки.
У высших растений содержатся важные классы регулирующих рост гормонов, основные из которых: ауксин, гиббереллины, цитокинины, абсцизовая кислота и этилен.
 Ауксин синтезируется растущими апикальными зонами стеблей, в том числе молодыми листьями. От апекса ауксин мигрирует в зону растяжения, где он специфически влияет на рост растяжением. Природный ауксин представляет собой простое соединение – индолил-3-уксусную кислоту (ИУК).
Ауксин синтезируется растущими апикальными зонами стеблей, в том числе молодыми листьями. От апекса ауксин мигрирует в зону растяжения, где он специфически влияет на рост растяжением. Природный ауксин представляет собой простое соединение – индолил-3-уксусную кислоту (ИУК).
Транспорт ИУК происходит полярно со скоростью 10-15 см/ч от вершины побегов к корням. Механизм полярного транспорта следующий: в апикальный конец клетки ИУК проникает пассивно совместно с Н+, а в базальном конце – активно секретируется через клеточную мембрану.
Физиологическое действие ауксина сложное. Разные ткани отвечают на действие ауксина увеличением роста, которое обусловлено стимуляцией растяжения клеток.
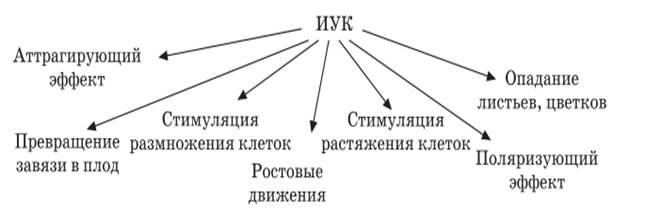
Аттрагирующее действие заключается в том, то клетки и ткани, обогащенные ауксином, становятся центрами притяжения веществ. Роль ауксина в стимуляции опадания листьев и цветов связана с заметным понижением его содержания в листьях. Это приводит к старению листьев. Стареющие ткани продуцируют этилен, который действует на отделительную зону (зона опадания).
Сам же ауксин задерживает опадание листа и цветов. Есть доказательства, что ауксин, кроме участия в растяжении клеток и опадании листьев, стимулирует процессы клеточного деления. Вероятно, ауксины повышают камбиальную активность. Считают, что ауксины участвуют в дифференциации сосудистой ткани в период начала ростового процесса и в образовании боковых корней. Превращение завязи в плод – еще один контролируемый ауксином процесс. Первостепенную роль играет ауксин в ростовых движениях – тропизмах и настиях.
В основе действия ауксинов существует два механизма: быстрое влияние ауксинов на мембранную систему, где за счет энергии АТФ увеличивается транспорт водородных ионов из цитоплазмы в клеточную оболочку и ускоряется размягчение клеточной оболочки; медленное влияние ауксинов через геномную систему на синтез белков, определяющих рост клеток.
Наличие обоих механизмов весьма вероятно, поскольку ауксин не только вызывает выход протонов, но и изменяет микроструктуру цитоплазмы (микротрубочки).
 Гиббереллины – фитогормоны, преимущественно класса тетрацикличес-ких дитерпеноидов. Все гиббереллины – карбоновые кислоты, поэтому их называют гибберелловыми кислотами. Известно более 110 разных гиббереллинов (ГК), многие из которых не обладают физиологической активностью в растениях.
Гиббереллины – фитогормоны, преимущественно класса тетрацикличес-ких дитерпеноидов. Все гиббереллины – карбоновые кислоты, поэтому их называют гибберелловыми кислотами. Известно более 110 разных гиббереллинов (ГК), многие из которых не обладают физиологической активностью в растениях.
ГК синтезируются главным образом в корнях и листьях. Транспорт их осуществляется пассивно с ксилемным и флоэмным потоками. ГК при нанесении на некоторые растения вызывают сильное удлинение стебля, а в ряде случаев и уменьшение листовой поверхности. Самое яркое проявление их действия – быстрая стимуляция удлинения цветоножки (стрелкование) и во многих случаях стимуляция цветения длиннодневных растений. У короткодневных растений ГК, вероятно, оказывают обратный эффект на цветение. Место действия ГК – апикальная и интеркалярная меристемы.
Обработка ГК выводит семена и клубни некоторых растений из состояния покоя. Экзогенно введенный ГК снимает у двухгодичных растений необходимость в яровизации, стратификации у тех семян, для которых стратификация необходима. ГК вызывают партенокарпию: для этого цветки необходимо опрыскивать раствором ГК. Ауксин тоже может вызывать партенокарпию, но ГК более активны. В тканях, обработанных ГК, увеличивается содержание ИУК. Считают, что физиологической основой карликовости большинства растений является нарушение гиббереллового обмена, который приводит к недостатку эндогенных гиббереллинов.

Ауксины и гиббереллины представляют два сильнодействующих класса регуляторов. Однако они не способны регулировать ход ростового процесса в онтогенезе.
 Цитокинины. Ни один из предыдущих гормонов не может повлиять на процесс зеленения изолированных листьев, закладку почек в культуре тканей. Этими свойствами обладают цитокинины, получившие свое название из-за способности стимулировать цитокинез (клеточное деление). Цитокинин обнаружен в некоторых бактериях, водорослях, грибах и насекомых. Основное место синтеза цитокининов – корни; однако в последнее время получены данные о том, что синтез цитокининов может происходить и в семенах. Из корней цитокинины пассивно транспортируются в наземные органы по ксилеме.
Цитокинины. Ни один из предыдущих гормонов не может повлиять на процесс зеленения изолированных листьев, закладку почек в культуре тканей. Этими свойствами обладают цитокинины, получившие свое название из-за способности стимулировать цитокинез (клеточное деление). Цитокинин обнаружен в некоторых бактериях, водорослях, грибах и насекомых. Основное место синтеза цитокининов – корни; однако в последнее время получены данные о том, что синтез цитокининов может происходить и в семенах. Из корней цитокинины пассивно транспортируются в наземные органы по ксилеме.

Роль цитокининов в процессах клеточных делений связана со стимуляцией репликации ДНК и регуляцией переходов из предшествующих фаз в фазу митоза. Есть данные о действии цитокинина на транспорт К+, Н+, Са2+.
Этилен (СН2 = СН2) – гормон старения (гормональный фактор газоподобного типа). Давно известно, что одно гнилое яблоко в бочке вызывает порчу всех остальных. Как оказалось, в гнилом яблоке вырабатывается летучее вещество – этилен, производящий разрушительное действие в здоровых плодах. Тот факт, что действие этилена можно снять повышенной концентрацией СО2 в окружающей среде, лежит в основе практического приема хранения яблок и других плодов.
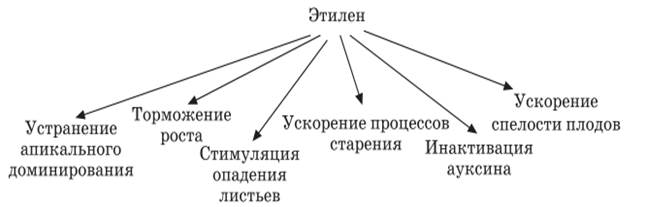
Этилен вызывает образование апикального изгиба во многих этиолированных проростках; действие света на выпрямление изгиба связано с тем, что свет ингибирует образование этилена. Этилен может влиять на геотропизм и другие опосредованные ауксином реакции (например, подавление роста боковых почек). Этилен тормозит полярный транспорт ауксина, усиливает процессы старения, опадения листьев и плодов, устраняет апикальное доминирование, а также ускоряет поспевание плодов.
 Абсцизовая кислота (АБК) – природный гормональный ингибитор роста терпеноидной природы.
Абсцизовая кислота (АБК) – природный гормональный ингибитор роста терпеноидной природы.
АБК синтезируется главным образом в листьях, а также в корневом чехлике. Перемещение АБК в растениях происходит как в базипетальном, так и в акропетальном направлениях в составе ксилемного и флоэмного соков.
Как мы уже отметили, в большинстве случаев АБК тормозит рост растений. Этот гормон чаще выступает как антагонист ИУК, цитокинина и гиббереллинов.

АБК ингибирует прорастание семян и рост почек, содействует опадению листьев, связанному с их старением. АБК ускоряет распад нуклеиновых кислот, белков и хлорофилла. В некоторых случаях АБК – активатор: стимулирует развитие партенокарпии у розы, удлинение гипокотиля огурца, образование корней у черенков фасоли. В высоких количествах АБК образуется при стрессах (при действии разных неблагоприятных факторов внешней среды). Особенно много ее образуется при водном стрессе в листьях. Действие АБК в этом случае обусловлено ее влиянием на функционирование Н+-помпы, вызывающим отток ионов К+ из замыкающих клеток, в результате чего устьица закрываются и тем самым предотвращается опасность высыхания. АБК может выполнять и сигнальную роль при водном дефиците.
Таким образом, АБК – ингибитор широкого действия, который влияет на процессы покоя, роста, движения устьиц, геотропизма, поступления веществ в клетку.
Фузикокцин был открыт как токсин, выделяемый патогенным грибом Phusicoccum amygdale. Широко распространен в природе. Сейчас известно более 15 соединений этой группы. Кроме грибов они содержатся в клетках водорослей, высших растений (моховидные, папоротникообразные, цветковые) и даже животных.
По химической природе фузикокцин относится к терпеноидам и у цветковых растений представляет собой дитерпен – С36Н56О12. По своему действию фузикокцин похож на ауксин. Он стимулирует растяжение клеток корней, стеблей, колеоптилей, листьев, причем даже активнее, чем ИУК, а также прорастание семян (например, пшеницы). Фузикокцин вызывает открывание устьиц в присутствии и отсутствие света (антагонист АБК). Это связано с активацией протонной АТФазы и калиевых каналов. Кроме того, фузикокцин стимулирует транспорт кальция, хлора, глюкозы, аминокислот в клетку, а также дыхание и корнеобразование.
Фузикокцин обладает антистрессовыми функциями. Он повышает всхожесть семян при повышенных и пониженных температурах, избыточном увлажнении, при засолении. Замачивание семян в растворе фузикокцина, а также опрыскивание им в фазу кущения озимых пшеницы, ржи и ячменя повышает их морозоустойчивость благодаря лучшему развитию фотосинтетического аппарата у обработанных растений и накоплению ими большего количества в клетках сахаров. Фузикокцин защищает растения риса при засолении, повышает устойчивость клубней картофеля к некоторым заболеваниям.
К группе фитогормонов относят также жасминовую и салициловую кислоты, а также обнаруженные в растениях в последние годы другие гормональные соединения. ЖК и ее метиловый эфир могут контролировать процессы созревания плодов, роста корня, изгиб усиков, производство жизнеспособной пыльцы, устойчивость растений к насекомым и патогенам. Жасминовая кислота и ее метиловый эфир могут синтезироваться при механическом повреждении из линоленовой кислоты, образующейся при распаде фосфолипидов клеточных мембран. Жасминовая кислота транспортируется в неповрежденные участки по флоэме, а ее этиловый эфир как летучее соединение через воздух действует на соседние растения, «информируя» их о нападении патогенов. Содержание ЖК в тканях растений возрастает при механических раздражениях: изменении тургорного давления при водном дефиците, движении усиков, взаимодействии корневых волосков с частицами почвы. Активация синтеза ЖК в ответ на ряд механических раздражений происходит с участием Са-кальмодулинового пути сигнальной трансдукции. Концентрация ЖК наиболее высока в зонах клеточного деления, молодых почках, цветках, тканях околоплодника, в гипокотильном крючке бобовых растений. ЖК снижает содержание хлорофилла и приводит к хлорозу.

Концентрация жасминовой кислоты в растительных тканях резко возрастает при механическом повреждении или воздействии элиситоров, способных индуцировать реакцию сверхчувствительности (СВЧ). Последняя – один из самых эффективных способов защиты растительных организмов от повреждения, поскольку происходит быстрая локальная гибель инфицированных растительных клеток вместе с патогеном, что и обеспечивает устойчивость всего растения. Синтез жасмонатов запускается при ранении растения, например в процессе поедания фитофагами.
Жасминовая кислота активирует экспрессию ряда генов, продукты которых вырабатываются в ответ на такие стрессовые воздействия, как механическое повреждение тканей и заражение патогенами. К их числу относятся, например, тионины, экстенсины, ферменты, участвующие в синтезе ряда фенольных соединений и фитоалексинов. Поэтому обработка растений жасминовой кислотой резко повышает их устойчивость к повреждениям. Жасминовая кислота – один из факторов индукции иммунитета растений к повторным заражениям.
Салициловая кислота также обеспечивает растению устойчивость к повреждениям, вызываемым различными патогенами. Синтез СК играет определяющую роль в реакции сверхчувствительности, а также в пролонгированной системной устойчивости растений к широкому кругу инфекций.
 Образование больших количеств пероксида водорода – также причина активации синтеза фитогормонов – салициловой и жасминовой кислот.
Образование больших количеств пероксида водорода – также причина активации синтеза фитогормонов – салициловой и жасминовой кислот.
Повышение содержания салициловой кислоты усиливает реакцию СВЧ, поскольку салицилат – ингибитор каталазы – фермента, расщепляющего пероксид водорода. То есть пероксид водорода, активируя синтез салициловой кислоты, способствует еще большему накоплению активных форм кислорода и вызывает усиление СВЧ-реакции. По мере затухания реакции СВЧ салициловая кислота переводится в связанную форму, взаимодействуя с глюкозой и образуя гликозиды. Именно салициловая кислота и образование ее конъюгантов – ключевые элементы не только в СВЧ-реакции, но и в формировании системного приобретенного иммунитета растения. Таким образом, в ходе СВЧ-реакции в растении происходит также формирование иммунитета к повторным заражениям.
Системин – полипептидный гормон, обнаруженный в растениях в 1991 г., состоит из 18 аминокислот. В отличие от ранее известных гормонов он запускает системы, защищающие растения от патогенов, повышает устойчивость к заболеваниям.
Важный компонент защитных реакций растений – ингибиторы ферментов, участвующие в процессах переваривания пищи насекомыми. Наиболее изучены из них ингибиторы протеолитических ферментов (протеазы), которые расщепляют белки в процессе пищеварения. У насекомых, которые питаются растениями, содержащими ингибиторы протеаз, резко падают темпы роста и развития, поскольку в их меню отсутствуют свободные аминокислоты. Ингибиторы протеаз обычно появляются в растениях в ответ на повреждение. Экспрессия генов, кодирующих ингибиторы протеаз, индуцируется механическим повреждением растений и непосредственно контролируется двумя фитогормонами: небольшим пептидом из 18 аминокислот, системином и жасминовой кислотой.
Показано, что синтез ингибиторов протеаз является результатом целого ряда событий.
На первом этапе повреждения растения насекомыми синтезируется системин – первый гормон пептидной природы, обнаруженный в растениях. Затем системин транспортируется по флоэме в неповрежденные участки растения, где взаимодействует с рецепторами и инициирует синтез еще одного гормона – жасминовой кислоты, которая в свою очередь активирует экспрессию генов, кодирующих синтез ингибиторов протеаз. При этом регуляция экспрессии некоторых «защитных» генов системином может осуществляться вместе с другими гормонами, например такими, как АБК, этилен, жасминовая кислота.
В растениях выявлены и другие гормоны, в частности фитосульфокины, участвующие в регуляции клеточных делений и других ростовых процессов у растений. Это небольшие пептиды, состоящие из 4-5 аминокислот.
В последние годы к регуляторам роста, проявляющим гормональную активность в растениях, стали относить брассиностероиды (БС) и простагландины (ПГ).
Негормональные регуляторы роста. Функции основных типов фитогормонов-стимуляторов (ауксины, гиббереллины, цитокинины) и их антагонистов – фитогормонов-ингибиторов (АБК и этилен) – реализуются в растительной клетке при непосредственном влиянии негормональных факторов. К последним относятся вещества, усиливающие действие фитогормонов (витамины, фенольные соединения (протекторы и синергисты), имитаторы фитогормонов).
Открытие фитогормонов предопределило предпосылки для создания химических основ ростовых веществ. Вообще, все природные и синтетические соединения, участвующие в регуляции роста и развития, объединяют под общим названием регуляторы роста и развития. К ним относятся органические соединения, отличные от питательных веществ, но вызывающие стимуляцию или ингибирование процессов роста и развития.
Природные стимуляторы роста. Витамины – негормональные регуляторы, синтезирующиеся в растениях в микроколичествах. Выполняют каталитические функции, усиливают ростовые процессы, активируемые фитогормонами. Фенольные протекторы фитогормонов – эндогенные соединения, предотвращающие разрушение или иммобилизацию регуляторных соединений. К ним относятся такие фенольные соединения, как фенолкарбоновые кислоты (кофейная, феруловая, синаповая), которые защищают ауксины типа ИУК от разрушения. Таким образом, протекторы являются непосредственными регуляторами метаболитов фитогормонов, т.е. регуляторами обмена фитогормонов. Фенольные синергисты – вещества, самостоятельно не участвующие в регуляции роста, но активирующие функцию фитогормонов. Как уже отмечалось, ни протекторам, ни синергистам самостоятельный гормональный эффект не свойственен. Этим они отличаются от имитаторов гормонов, т.е. органических соединений бактериального, грибного или растительного происхождения, которые проявляют на биотестах эффект одного или нескольких гормонов растений.
Природные ингибиторы роста. Соединения негормональной природы представляют собой самостоятельный класс регуляторов в растениях. Сюда относят ряд фенольных производных (паракумаровая кислота, флоридзин и др.), а также терпеноидные соединения (портулал, кукурбитацин, бататазины и др.).
Синтетические регуляторы роста. За последние годы был получен ряд синтетических регуляторов, в частности синтетических ингибиторов. Последние соединения составляют несколько групп, владеющих специфичными функциями, например:
1. Реторданты, подавляющие рост стебля (хлорхолинхлорид, фосфон и др.);
2. Антиауксины, тормозящие перемещение ИУК и ее аналогов по растению.
3. Гербициды – синтетические препараты, убивающие растения. Рассматривают гербициды общего действия, которые уничтожают всю растительность, селективные – для борьбы с сорняками в монокультурах. При действии гербицидов сначала нарушается полярность, утолщаются побеги, появляются эпинастии, опадают листья, наступают паталогические морфозы, что и приводит к гибели растения.
Химические аналоги природных ингибиторов. Это синтетические соединения, аналоги этилена, фенольных ингибиторов, кумаринов и АБК, владеющие мощным гербицидным или дефолирующим действием.
В последние годы большое распространение получили препараты, усиливающие цветение плодовых культур (алар), вызывающие дефолиацию и задержку роста стебля (этрел, гидрел).
Химические аналоги природных стимуляторов роста. Это регуляторы, активирующие особые фазы роста и онтогенеза растений. К ним, например, относятся синтетические аналоги ауксинов (а-нафтилуксусная, идолилмасляная, 2,4-дихлорфеноксиуксусная кислоты). Стимуляторы роста применяют для активации корнеобразования, роста культуры тканей, предотвращения опадения плодов.