1 Клеточные основы роста
2 Особенности роста органов растений
КЛЕТОЧНЫЕ ОСНОВЫРОСТА
Типы роста у растений. У многоклеточных растений в отличие от животных рост (за исключением ранних стадий развития зародыша) происходит только в определенных участках, меристемах.
Меристемы – это зоны в растительном организме, где происходит регулярное размножение растительных клеток. Эти зоны расположены апикалъно, т.е. на вершине растущего органа (в главных и боковых побегах и корнях), базипетально (в листьях и междоузлиях) или интеркалярно, например над узлами в соломине злаков. Интеркалярная – это вставочная меристема. Между листом и стеблем в пазухах листьев закладываются пазушные почки. Пазушные почки, которые длительное время не дают побегов, называют спящими; при определенных условиях они пробуждаются и из них развиваются побеги. Латеральная (камбий) меристема лежит параллельно вдоль длинной оси органа (например, пробковый камбий) и обеспечивает утолщение. Внутренние физиолого-биохимические реакции координируют ход ростового процесса на всех этапах жизни, определяют механизмы роста. Различают первичные и вторичные механизмы роста.

К первичным относят физиолого-биохимические реакции, лежащие в основе начальных этапов ростового процесса (лаг-фаза) и фазы ускоренного роста (логарифмическая фаза). К этим же механизмам относят электрофизиологические, гормональные и генетические реакции, которые запускают и поддерживают нормальный ход роста клеток, тканей и органов.
Вторичные механизмы роста – физиолого-биохимические реакции, участвующие в нормальном ходе роста (лаг-фаза и фаза замедления роста) и происходящие в процессе жизнедеятельности растений. К ним относят корреляцию между органами, донорно-акцепторные связи, метаболическую координацию между ростом и другими физиологическими процессами (фотосинтез, транспорт, запасание веществ, стресс).
Таким образом, можно выделить два типа роста: первичный и вторичный. В результате первичного роста может образоваться целое растение (для большинства однолетних и травянистых двудольных это единственный тип роста). В нем участвует апикальная, а иногда и интеркалярная меристемы. У некоторых растений за первичным ростом идет вторичный рост, в котором участвуют латеральные меристемы. Такой тип роста характерен для кустарников и деревьев. У ряда травянистых растений наблюдается вторичное утолщение стебля, например развитие дополнительных проводящих пучков у подсолнечника. Выделяют еще диффузионный рост (рост во время деления всех клеток).
Клеточные основы роста. Основой роста является образование новых клеток и их рост. Новые клетки образуются в результате деления клеток меристемы. Онтогенез клетки – это период от возникновения клетки до ее смерти. Физиологическая основа роста на каждой фазе различна.
Эмбриональная фаза. Клетка начинает свое существование благодаря делению клетки меристематической ткани. На этой фазе клетки любого органа и любого растения похожи друг на друга. Они мелкие, изодиаметрические, имеют тонкие клеточные стенки и цитоплазму с крупными ядром и ядрышком. После деления начинается рост ядра и цитоплазмы, в результате каждая дочерняя клетка вырастает до размеров материнской. При этом количество всех органелл и молекул таких веществ, как ДНК, РНК, белки, липиды и прочие, удваивается, т.е. восстанавливается их первоначальное количество, бывшее в материнской клетке до ее деления. На этой фазе многие органеллы представлены своими предшественниками: промитохондриями, пропласти-дами, провакуолями. В это время в клетках существуют два типа митохондрий: крупные гантелевидные и мелкие с разной степенью развития крист. Эндоплазматический ретикулум агранулярный, в цитозоле много свободных рибосом. Таким образом, во время эмбриональной фазы сама клетка растет очень медленно, но в это время быстро идет новообразование элементов структуры (органелл), формируется первичная клеточная стенка.
Во время этой фазы питательные вещества, поступающие в клетку, используются в основном для образования органелл или превращаются в запасные соединения. Распад белков замедлен; разнообразие ферментов меньше, чем во время других фаз.
Величина дыхательного коэффициента больше единицы, так как в клетках интенсивно идут не только аэробные, но и анаэробные процессы. Такой тип дыхания выработался, очевидно, как адаптация к гипоксии, которая часто возникает в местах расположения меристематических тканей, особенно в почках. Благодаря высокой интенсивности гликолиза в клетке образуется много промежуточных продуктов окисления, необходимых для быстрого синтеза органических веществ, в первую очередь, нуклеиновых кислот и белков. Предполагают, что менее активный окислительный обмен благоприятствует делению клеток. Движение цитоплазмы замедленно, вязкость увеличена.
После достижения размера материнской клетки новая клетка может снова разделиться и таким образом остаться на эмбриональной фазе или перейти в фазу растяжения. Клетка, остающаяся на эмбриональной фазе, может еще делиться 5-6 раз, после чего тоже растягивается.
Фаза растяжения. На этой фазе клетка быстро растет в длину, т.е. растягивается. Отсюда и название фазы.
Рост растяжением – это очень быстрый тип роста, свойственный только растительным клеткам. Впервые он появился у нитчатых водорослей. Клетка, имевшая размер 5-10 мкм, увеличивается в 10-50 и даже в 100 раз со скоростью до 100 % в час. Клетки мякоти околоплодника тыквы могут увеличиться на этой фазе в 1 млн раз. Распускание почек весной или после сухого сезона происходит в основном в результате быстрого растяжения клеток.
Рост растяжением делят на несколько этапов: подготовка к растяжению, собственно растяжение, фиксация объема клетки, торможение и остановка роста.
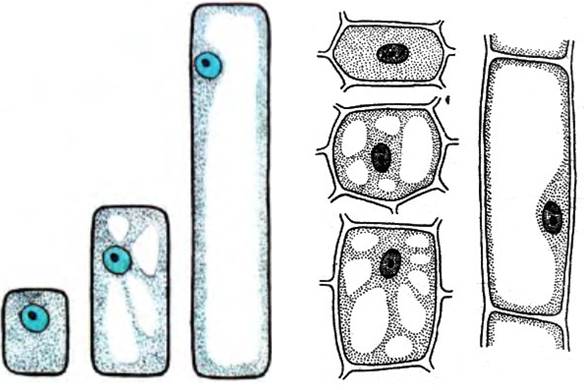
На первом этапе в клетке, готовящейся к растяжению, расширяются цистерны эндоплазматического ретикулума, из которых возникают вакуоли; свободные рибосомы образуют полирибосомы или прикрепляются к мембранам ЭПР; быстрее синтезируются белки, пектиновые вещества, РНК; увеличивается отношение РНК/белок; накапливается крахмал; увеличивается интенсивность дыхания и число митохондрий.
Второй этап – собственно растяжение – связан прежде всего с образованием крупной центральной вакуоли. Из расширяющихся цистерн эндоплазматического ретикулума возникают маленькие вакуоли. При этом тонопласт начинает захватывать с помощью эндоцитоза старые митохондрии, кусочки цитоплазмы, крупные молекулы. Затем с образовавшимися вакуолями соединяются лизосомы, содержащие гидролитические ферменты. Крупные молекулы, остатки органелл под действием этих ферментов разрушаются. В результате в вакуолях увеличивается концентрация осмотически активных веществ и в них начинает поступать вода. Количество воды в растягивающихся клетках, например корня, увеличивается в 7-35 раз. Вакуоли быстро увеличиваются и, наконец, сливаются друг с другом, образуя одну или несколько центральных вакуолей, а цитоплазма располагается тонким слоем вокруг них. Обычно объем вакуоли растет быстрее объема цитоплазмы.
Еще до образования центральной вакуоли происходит разрыхление и растяжение клеточной стенки. Вакуоль давят на цитоплазму, и клетка растягивается. Тургорнос давление – главная сила, растягивающая клетку. Чтобы клеточная стенка могла растягиваться, у нее должна увеличиться эластичность. Увеличение эластичности клеточной стенки происходит в результате разрыва с помощью ферментов химических связей между целлюлозой и пектиновыми веществами, увеличения секреции белка клеточной стенки – экстенсина, уменьшения количества кальция в пектатах (выделения кальция).
Таким образом, в фазе растяжения рост клетки складывается из тесно связанных друг с другом нескольких процессов: продолжающегося увеличения количества цитоплазмы, увеличения объема клеточного сока и соответственно вакуоли, увеличения массы клеточной стенки, синтеза макромолекул.
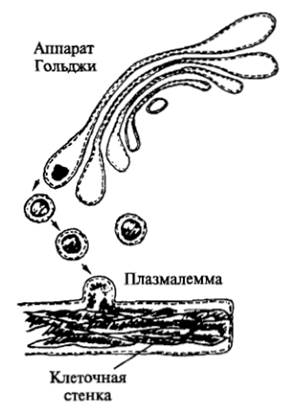 На третьем этапе, после того как растяжение произошло, объем клетки надо зафиксировать. Как происходит эта фиксация? Восстанавливаются ранее разорвавшиеся химические связи между молекулами целлюлозы и пектиновых веществ. Идет интенсивное образование микрофибрилл целлюлозы. Для этого активность аппарата Гольджи усиливается, его везикулы приносят необходимые ферменты, а также гемицеллюлозы, пектиновые вещества и, возможно, гликопротеиды к плазмалемме.
На третьем этапе, после того как растяжение произошло, объем клетки надо зафиксировать. Как происходит эта фиксация? Восстанавливаются ранее разорвавшиеся химические связи между молекулами целлюлозы и пектиновых веществ. Идет интенсивное образование микрофибрилл целлюлозы. Для этого активность аппарата Гольджи усиливается, его везикулы приносят необходимые ферменты, а также гемицеллюлозы, пектиновые вещества и, возможно, гликопротеиды к плазмалемме.
В результате экзоцитоза содержимое везикул попадает в периплазматическое пространство. Новые микрофибриллы целлюлозы синтезируются у наружной поверхности плазмалеммы и встраиваются в клеточную стенку между уже имеющимися микрофибриллами. Мембраны везикул, включаясь в плазмалемму, увеличивают ее.
На четвертом этапе изменяется соотношение гормонов, что приводит к торможению растяжения клетки.
Растяжение клетки сопровождается увеличением количества и усложнением строения органелл. Резко увеличивается количество диктиосом и везикул, что связано с высокой секреторной активностью, необходимой для образования клеточной стенки. Эндоплазматический ретикулум в это время уже хорошо развит, так как участвует в образовании вакуолей. Как уже говорилось, увеличивается количество рибосом, которые связываются с мембранами ЭПР или образуют полирибосомы; много амилопластов. Количество митохондрий увеличивается в 5 раз. Происходит деление митохондрий, пластид, а также их дифференцировка.
В растягивающейся клетке резко увеличивается количество не только воды, но и сухого вещества. Во время растяжения синтезируются вещества клеточной стенки, а также белки и липиды. В клетках бобовых иногда происходит даже удвоение количества белков. Однако преобладают в растягивающихся клетках небелковые соединения азота: свободные аминокислоты, нуклеотиды и др. Увеличивается количество нуклеиновых кислот (РНК), что способствует активации белкового синтеза в начале растяжения. Далее в течение растяжения содержание белка и РНК в пересчете на клетку, как правило, уменьшается. Изменяется углеводный обмен. Общее количество сахаров выше, чем в меристематических клетках. В растягивающихся клетках, например корня, вместо сахарозы, преобладающей на эмбриональной фазе, накапливаются моносахара (глюкоза, фруктоза). Их количество увеличивается в 6-7 раз. Доля целлюлозы увеличивается до 30 % и более. Количество низкомолекулярных веществ (сахаров, органических кислот, неорганических ионов и др.) в клетке увеличивается быстрее, чем
высокомолекулярных. В 10 раз увеличивается количество пирувата, фосфоенолпирувата и глюкозо-6-фосфата.
На химический состав клеток влияет их расположение. Так, растягивающиеся клетки колеоптиля содержат больше свободных аминокислот, чем растягивающиеся клетки корня.
Активность некоторых ферментов резко увеличивается по сравнению с эмбриональной фазой. Например, активность нитратредуктазы и глутаматдегидрогеназы увеличивается в корнях кукурузы в 10-50 раз. Возрастает активность гидролитических ферментов, что связано с образованием вакуолей.
Накопление низкомолекулярных веществ стимулирует дыхание. Основной путь дыхания во время этой фазы роста – гликолиз и цикл Кребса. Усиливается также пентозофосфатный окислительный путь, который поставляет НДДФН и рибозы для синтеза нуклеиновых кислот. Усиление дыхания поддерживается как увеличением числа митохондрий в клетке, так и достаточным количеством дыхательного субстрата и увеличением активности гликолитических ферментов. Главным дыхательным субстратом являются сахара, поэтому дыхательный коэффициент равен 1.
Обладающая максимальной активностью аэробного дыхания, максимальной скоростью роста, интенсивным обменом веществ, клетка в фазе растяжения минимально устойчива. В результате в неблагоприятных условиях в первую очередь повреждаются клетки, находящиеся в фазе растяжения.
Фаза дифференцировки клетки. Дифференцировка – это возникновение структурных и функциональных различий между клетками, а значит, между тканями и органами в процессе развития растения. В результате дифференцировки растянувшиеся клетки превращаются в специализированные в строгом соответствии с функцией гой ткани, в состав которой данная клетка будет входить. Существует несколько типов дифференцировки.
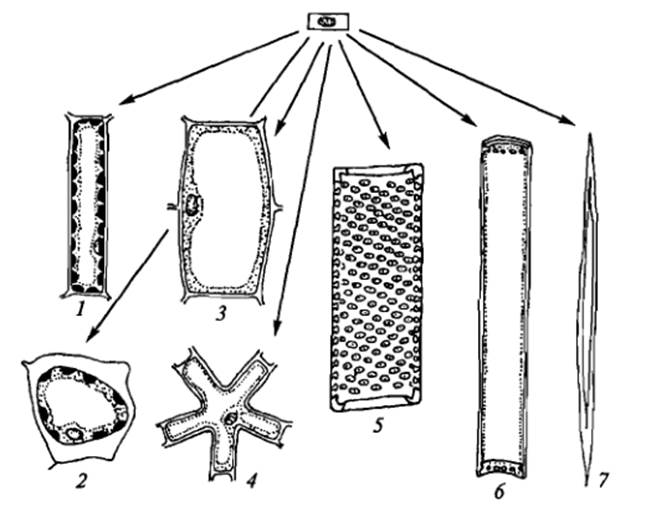
Дифференцировка клеток:
1– палисадная; 2– замыкающая; 3 – паренхимная; 4 – звездчатая; 5 – членик сосуда; 6 – членик ситовидной трубки; 7 – склеренхимное волокно
Структурная (морфологическая) дифференцировка – это возникновение различий по морфологическим признакам. На клеточном уровне она выражается в различной толщине и структуре клеточной стенки, форме клеток, разной степени их вакуолизации, особенностях развития тех или иных органелл.
Биохимическая дифференцировка – это возникновение различий в составе ферментов, в способности к синтезу запасных веществ или вторичных метаболитов и в других изменениях в клетке, влияющих на обмен веществ.
Физиологическая (функциональная) дифференцировка – это формирование между клетками различий, приводящих к выполнению ими разных функций, например, различий между авто- и гетеротрофными клетками, соматическими клетками и гаметами, между клетками разных тканей. Биохимическая дифференцировка предшествует другим ее типам.
В фазе дифференцировки возникает многообразие клеток. При этом может происходить как усложнение, так и упрощение структуры клетки. В процессе дифференцировки у одной клетки разрушаются поперечные клеточные стенки и утолщаются за счет отложения лигнина продольные, разрушается протопласт, и она превращается в членик сосуда. У другой клетки исчезает клеточное ядро и тонопласт, и она превращается в членик ситовидной трубки. В течение дифференцировки количество митохондрий в клетке может сильно увеличиться (образование железистой клетки или сопровождающей клетки во флоэме) или уменьшиться (образование эпидермальной клетки), могут образоваться хлоро-, хромо или лейкопласты или огромная вакуоль и боковой вырост, в который передвигается ядро (образование корневого волоска).
Таким образом, после окончания растяжения одинаковые до сих пор клетки начинают развиваться по-разному. Приобретение клеткой, тканью, органом, организмом способности реализовать определенные наследственные признаки называется детерминацией. Детерминация характеризуется возникновением способности к развитию по определенному пути с одновременным ограничением возможностей развития в других направлениях.
Почему растянувшаяся клетка может превратиться в клетку другой ткани? Направление дифференцировки зависит от набора ферментов в клетке, а синтез каждого фермента регулируется генами. Все клетки организма происходят из зиготы в результате ее деления. Зигота обладает полным набором генов. Митозы, как правило, являются равнонаследственными делениями, поэтому новые клетки имеют такой же набор генов, как и материнская клетка, обладают одинаковыми потенциальными возможностями (тотипотентностью), т.е. способны синтезировать все белки данного организма. Однако не все гены активны одновременно, лишь часть существующих генов активна, поэтому каждая клетка синтезирует лишь определенный набор ферментов, реализуя лишь часть своих генетических возможностей. В результате клетки одного многоклеточного организма сильно отличаются по форме и функциям, например клетки листа и корня, флоэмы и ксилемы.
Активация (дерепрессия) одной группы генов при одновременной инактивации (репрессии) другой (дифференциальная активность генов) приводит к образованию различных наборов ферментов в разных клетках. Эта биохимическая дифференцировка является причиной структурной дифференцировки. В процессе дифференцировки клетки не происходит ни потери, ни приобретения новых генов, изменяется только их активность. В течение жизни каждой клетки реализуются только некоторые из этих потенций.
Итак, разные клетки многоклеточного организма, различающиеся по форме и функциям, отличаются по составу ферментов.
На активность генов влияют разные факторы, например местоположение клетки. Если дифференцирующаяся клетка находится на поверхности органа, то она превращается в клетку покровной ткани, если в центре, – то основной (стебель) или проводящей (корень).
Другим важным фактором являются контакты между клетками. У многоклеточных растений клетки в тканях и органах связаны друг с другом плазмодесмами, транспортирующимися веществами. Любые пространственные или временное изменения этих связей влияют на дифференцировку. Изоляция неделящейся клетки, лишение ее связи с соседними клетками приводит к превращению ее в делящуюся клетку, дающую начало каллусу или зародышу. Каллус – ткань, возникающая у растений в местах повреждений, существующая недолго и способствующая заживлению ран. Метод выращивания на искусственной питательной среде в стерильных условиях клеток и тканей, возникших в результате деления клеток, выделенных из кусочков листа, стебля, корня или других органов, называется культурой изолированных клеток и тканей. В культуре изолированных клеток и тканей рост клеток состоит из этих же фаз, но эмбриональная фаза очень длинная. Если в норме клетка делится 5 раз, то в культуре может делиться много лет, не переходя к растяжению.
Фактором дифференцировки является и способ деления клетки: равное или неравное деление. При неравном делении поверхностной клетки корня или листа из меньшей дочерней клетки соответственно образуется корневой волосок или замыкающая клетка устьица, а из большей – клетка эпиблемы или эпидермы. В культуре изолированных клеток в результате равного деления микроспоры образуется каллус, а неравного – зародыш.
В течение эволюции растений дифференцировка шла от небольшого числа дифференцированных клеток (2-3 типа у нитчатых водорослей) до все более сложных многоклеточных структур. У покрытосеменных растений 6 типов тканей: меристематические, покровные, проводящие, механические, секреторные, основные, которые еще можно разделить на 74-80 подтипов.
Обмен веществ в дифференцирующейся клетке характеризуется высокой скоростью дыхания, интенсивным синтезом ферментов и других белков, нужных для выполнения ее функции. Рост клетки на этой фазе замедляется и затем прекращается.
Дифференцированная клетка может вернуться к эмбриональной активности. Например, если у листа бегонии надрезать крупные проводящие пучки и положить его на влажный песок, то скоро образуются новые побеги, из которых можно получить взрослые растения. Новый побег возникает из одной эпидермальной клетки, которая делится на две, потом каждая дочерняя клетка вновь делится. Так на месте старой клетки возникает новая меристема, дающая начало побегу.
Чтобы специализированные клетки начали опять делиться, они должны дедифференцироваться. Дедифференцировка – это переход специализированных неделящихся клеток к делению, т.е. восстановление меристематической активности. Дедифференцировка, как процесс обратный дифференцировке, состоит в следующем: некоторые органеллы, например хлоропласты, разрушаются; клеточная стенка разрыхляется и набухает; увеличивается количество элементов аппарата Гольджи и эндоплазматического ретикулума.
Одновременно изменяется активность генов и идет подготовка аппарата белкового синтеза: увеличивается число ядрышек, происходит новообразование РНК; образуются полирибосомы. В результате появляются новые белки, свойственные делящимся клеткам. Эти преобразования происходят в основном за счет запасных веществ. Возникает меристемоподобная клетка, способная к делению. Во время дедифференцировки хорошо обнаруживается тотипотентность клетки. Дедифференцировка связана с активацией ранее не работавших генов.
Дедифференцировка происходит и при образовании раневого каллуса. Ближайшие к поверхности разреза паренхимные клетки сначала увеличиваются в размере, затем у них возобновляется рост цитоплазмы и, наконец, начинаются митозы. Образуется масса недифференцированных клеток, закрывающих рану (каллус). Из каллуса позднее в результате новой дифференцировки (редифференцировки) возникают различные ткани. У древесных растений каллус образуется преимущественно из камбия. Повреждение листовой пластинки может вызвать деление клеток мезофилла и формирование опробковевшей защитной ткани.
Не все клетки могут дедифференцироваться. Особенно легко дедифференцировка происходит у паренхимных, эпидермальных, железистых, сопровождающих клеток. Членик ситовидной трубки, клетки колленхимы, механических волокон не могут дедифференцироваться, так как у них произошли сильные изменения в процессе их собственной дифференцировки.
Таким образом, при вегетативном размножении, травмах происходит передифференцировка клеток. Например, при укоренении стеблевых черенков клетки нижней части стебля дедиффсренцируются, начинают делиться и образуют клетки придаточного корня. В основе передифференцировки лежит изменение активности генов.
Итак, на каждой фазе происходит рост клетки и новообразование ее структур, значительное же увеличение размеров самой клетки характерно только для фазы растяжения. Морфологически дифференцировка заканчивается достижением клеткой ее окончательного размера.
Деление процесса роста клетки на фазы относительно. Например, дифференцировка существует уже у клеток, находящихся на эмбриональной фазе. Они отличаются от других клеток меристемы биохимически и структурно: имеют меньше РНК, белка; у них хуже развит аппарат Гольджи, эндоплазматический ретикулум. Однако на фазе дифференцировки морфологические различия между клетками лучше выражены и видны в световой микроскоп. Кроме того, эти фазы онтогенеза клетки идут последовательно только в корне, стебле, колеоптиле. В листьях двудольных растений растягивающиеся клетки продолжают делиться, а некоторые из них одновременно дифференцируются. Обычно после окончания дифференцировки клетка приступает к выполнению специфической функции. Однако в листе активное функционирование совпадает с растяжением. Не все клетки после эмбриональной фазы растягиваются. Некоторые из них опять делятся, а другие переходят в состояние покоя. Причины перехода клетки на один из этих путей пока не ясны.