Предположение о том, что поведением ранних homo могли управлять деменцииe, галлюцинозы, мании, стереотипии, кататонияf и эхолалияg и так далее, т.е. то, из чего «состоит» (по И. Павлову) более-менее полноценная шизофрения в её сегодняшнем понимании, конечно, экзотично, но, полагаю, может иметь место лишь в романах, «житиях» или психологических сочинениях. Нейрофизиология же не оставляет сомнений в том, что только абсолютно адекватная и предельно точная рефлексия могла обеспечить выживаемость организму (виду, роду, семейству, отряду и так далее).
e Приобретённое слабоумие. — Прим. ред.
f Двигательные расстройства. — Прим. ред.
g Неконтролируемое повторение слов, услышанных в чужой речи. — Прим. ред.
Но следует признать, что общая стилистика первобытного мышления человека при поверхностном анализе даёт основания признать некой нормой этого мышления то, что в XIX столетии получило название «шизофрения». Нет сомнения, что абстрагированная психиатрическая диагностика XIX-XXI веков именно так маркировала бы и ацтекский способ приумножения урожая тыкв, а также принятые в средневековой Тюрингии методы укрепления стен через замуровывание в них детей, или эвхаристические практики. Но основываясь на сделанном выше анализе, мы получаем право на вывод, что эти и другие системные прецеденты истории homo не имеют ничего общего с деменциями или иными повреждениями функций мозга, а есть, вероятно, следствие несопоставимости поставленных жизнью задач и возможностей «природного» разума человека. К тому же, шизофрения — явление комплектное, и кроме алогичного ответа на сложный «вызов» предполагает наличие у «ответчика» множества иной симптоматики, а данными о таковой ни классическая антропология, ни история не располагают. Более того, люди, массово демонстрирующие неадекватность при решении сложных вопросов, показывают точность, успешность, хорошую обучаемость в решении тех задач, где все составляющие очевидны. (В вопросах строительства, ремёсел, военного дела, иерархирования своих сообществ и так далее.)
Исходя из вышесказанного разум не надо переоценивать, в нём нет ничего чрезвычайного или «уникального», это очень древняя и весьма банальная функция практически любого живого организма. Перечисленные выше факты и выводы также дают основания предполагать, что разум не является «главной функцией мозга».
Возникает два итоговых вопроса, касающиеся природы разума. Первый — о степени его развитости у homo сравнительно с другими животными.
Второй — насколько анатомическая сложность мозга может служить индикатором этой развитости?
На первый вопрос, ответ, полагаю, прост и очевиден: если развитость разума определять по точности тех поведенческих решений, что основаны на оценке явных факторов, то тут человек будет примерно равен любому другому животному, что подтверждают два миллиона лет его «дикой» истории и задокументированные образчики поведения «мауглеоидов».
К слову, теперь становится отчасти понятно, почему на простую задачу по усовершенствованию каменных орудий человеку понадобилось около миллиона лет.
В тех же случаях, когда поставленный перед «чистым разумом» вопрос содержит скрытые или неявные факторы, то основываясь лишь «сам на себе», отделённый от искусственных коллективных интеллектуальных систем, разум homo способен к решениям либо полностью ошибочным, либо содержащим очень высокий «процент ошибки», что в известной степени доказывают примеры, предложенные классической антропологией, религиями, народной медициной и так далее.
Что же касается второго вопроса, то однозначный ответ на него, вероятно, невозможен по простой причине отсутствия анатомического «субстрата разума».
Установив специализации практически всех отделов головного мозга, от мозжечка до лучистостей мозолистого тела, нейрофизиология не может и, вероятно, не сможет никогда указать ту мозговую структуру, которая собственно и генерирует интересующую нас функцию, т.е. разум.
К примеру:
Мы можем с определенной уверенностью не просто назвать те мозговые субстраты, которые осуществляют наш процесс визуализации мира, но и предъявить их «на стекле».
Для этого достаточно положить анатомический препарат головы homo или любого другого млекопитающего на стол — и отпрепарировать из него глаза, т.е. внешнюю часть зрительного анализатора головного мозга. Затем, вскрыв череп, достав и перевернув мозг, следует отделить оптический нерв, хиазму, зрительные тракты, прорезаться к латеральным коленчатым телам, лучистости Грациоле, клину, шпорной борозде, язычной извилинке, а затем коротким движением ланцета отодвинуть извлеченные структуры от основной мозговой массы.
Этот набор анатомических фрагментиков и будет материальным субстратом нашего зрения.
К слову, разумеется, это очень упрощённая, грубая схема препаровки, так как для анатомической комплектности зрительного анализатора потребуются сверхтонкие манипуляции по отделению зрачковых иннерваторов, чёрного вещества, ядер Вестфаля — Эдингера, Якубовича, протектального, красного ядер и так далее. Можно секционировать ещё эффектнее: вслед за отпрепаровкой глаз в орбите, сделать выпилку части большого крыла клиновидной кости и глазничной части лобной кости, а потом, после открытия черепа и мягкой оттяжки мозга от лобно-лицевой части, просто втянуть внутрь глаза, не нарушая их связи с оптикусом, хиазмой и зрительным трактом.
Мы сможем попытаться анализировать его по прямым признакам развитости (например, по количеству нервных волокон в n. opticus или в radiatio optica) и отчасти оценить его развитость и сложность.
При этом мы обязаны будем признать, что «потеряли» в ходе нашей препарации самое главное и ценное — т.е. те связи зрительного тракта со стволовыми ретикулярными и прочими структурами мозга, что побуждали его функционировать, и «пользовались» результатами его деятельности.
К слову, эти связи не всегда прямы; к примеру, связь с лимбической системой и гипоталамусом идёт через медиальные ядра таламуса.
Тем не менее пусть и лишенный движущих его связей, перед нами, зримый и осязаемый, будет лежать анатомический субстрат функции зрения.
Примерно такую же процедуру мы можем проделать практически с любой функцией головного мозга: со слухом, обонянием, тактилизацией, болью, моториками, хеморецепцией (вкусом), вокализацией и так далее.
Естественно, «набор анатомических фрагментиков» каждый раз будет существенно отличаться и количеством и размером отпрепаровок, не говоря уже о тех случаях, когда к нашему маленькому собранию «на стекле» придётся добавлять бесконечные ветвления висцеральной нервной системы, извлечённой из внутренних органов.
Ничего подобного с функцией разума мы проделать не сможем. Эта функция не имеет своего собственного субстрата, а является результатом взаимоотношений ретикулярной формации, гиппокампа и иных стволовых древних структур с анализаторами и рецепторами.
Констатация этого обстоятельства не подразумевает никакого драматического подтекста, она всего лишь выводит нас на вывод, что пытаться искать индикацию развитости разума имеет смысл лишь в простом наличии связей и трактов, а вот общее развитие головного мозга таким индикатором являться, по всей вероятности, не будет.
Да, увеличение общего объёма мозга и усложнение корковых структур, обилие ассоциативных и проекционных зон коры головного мозга, скорость и стабильность движения медиаторов — всё это является несомненным свидетельством ароморфоза. Но как демонстрируют приведённые выше примеры из народной медицины, религий, мифологии, к качеству «основных продуктов», т.е. ответов на сложные вопросы, поставленных средой, всё это не имеет ни малейшего отношения.
Если бы разум был полноценной, многообъемлющей функцией, которая способна предложить эти «верные ответы», то никакой потребности в интеллекте, вероятно, никогда бы и не возникло, как не возникло бы потребности в каменном рубиле, если бы зубы и когти homo были бы в состоянии сами вспороть кожу на приготовленной к съедению туше.
Иными словами, все впечатляющие современные возможности homo не имеют к разуму никакого отношения, а связаны исключительно с интеллектом, с искусственно созданной поколениями системой обмена и хранения относительно достоверной информации, образовавшейся и развившейся через конфликты опыта с традициями, мифами, религиями и другими порождениями чистого разума («первобытным мышлением»).
Вычислить точную точку отсчёта, некий момент зарождения этой системы не представляется возможным, но, по всей вероятности, она возникла практически одновременно с первыми глобальными экзерсисами первобытного мышления и сосуществовала с ними параллельно, то конфликтуя, то контактируя, но никогда не «сливаясь». Первоначально представленная одиночками, решившимися на конфликт с традицией и нормой, она долгое время существовала в «тени» чистого разума и не имела особого влияния. Порождена была, по всей вероятности, простыми агрессиями (praedonia, justa), желанием получить преимущества через обладание верными ответами на сложные вопросы и наблюдениями за неэффективностью тех решений, что предлагало первобытное мышление. До накопления того опыта, который мы можем сегодня маркировать как «естественнонаучный», была неэффективна, порождая лишь размышления, подобные тем, что прославили Фалеса Милетского или милые нелепости Аристотеля. Отчасти, «веками рождения» интеллекта (полного антагониста мышления первобытного) можно считать времена появления космогоний Анаксимандра, Ферекида Сиросского и Анаксимена. Впрочем, учитывая, что независимость их разработок от древнеиранской космогонии остается дискутивной и трудноопределяемой даже для крупнейших авторитетов в этой зыбкой области (таких как М. Л. Уэст, Э. Целлер, Э. Рет, Г. Дильс), «точная дата» зарождения интеллектуальной системы навсегда останется неизвестной. (Ignoramus et ignorabimus. — Не знаем и не узнаем.)
Исходные, базовые способности разума homo очень не велики и, вероятно, не превышают таковых у большинства животных с развитыми связями структур мозга. (Подтверждением этого тезиса служит палеолитическая ипостась человека, когда тот в течение бесконечно долгого периода являлся самым обычным животным.)
Определённые преимущества, позволившие человеку сделать столь удачную эволюционную карьеру, т.е. создать орудия, первоначальные механизмы, ремёсла, письменность, строительные навыки, по всей вероятности, бесполезно искать лишь в особенностях строения и объёма его мозга.
Но эти преимущества, несомненно, есть, следовательно, есть вопрос, требующий ответа.
Традиции естествознания в целом и нейрофизиологии в частности предлагают в качестве области поиска этих преимуществ внецеребральные анатомические особенности homo и прежде всего — его передние конечности:
«Нервная система, как инициатор в овладении приматами сложными условиями древесной жизни, привела к формированию руки. При переходе в эволюции человека от древесной жизни к прямохождению на ногах, руки стали не органами движения и передвижения, а органами метания и труда....Удачное сочетание центра (мозга) и периферии (рук), “инициатива” мозга и активность в овладении и использовании сложных условий древесной жизни стали фактором чрезвычайной важности в эволюции человека» (Стрельников И. Д. Анатомо-физиологические основы видообразования позвоночных, 1970).
Ещё более категоричен Г. Шеперд: «Главный интерес для нас представляют передние конечности, так как из них развились рука и кисть руки, т.е. органы, имевшие особое значение в эволюции приматов. Кисть руки — одно из наиболее эффективных и универсальных приспособлений, возникших в процессе филогенеза. Манипуляционные возможности кистей рук сыграли решающую роль в эволюции приматов» (Шеперд Г. Нейробиология, 1987. Т. 2).
Следует отметить, что данное понимание не является новацией или открытием физиологии XX века, оно с полной отчётливостью было сформулировано ещё Клодом Адрианом Гельвецием в XVIII столетии: «Если бы природа создала бы на конце нашей руки не кисть с гибкими пальцами, а лошадиное копыто, тогда, без сомнения, люди не знали бы ни ремёсел, ни жилищ, не умели бы защищаться от животных, и озабоченные исключительно добыванием пищи и стремлением избежать диких зверей, всё ещё бродили бы в лесах пугливыми стадами» (Гельвеций К. А. Об уме, 1973).
Полагаю, из трёх предложенных выше реплик о роли кисти руки в эволюционной судьбе человека, наиболее содержательной и точной является реплика К. А. Гельвеция, т.к. в её романтическом категоризме больше подлинной физиологии, чем в аккуратных выводах академиков XX века.
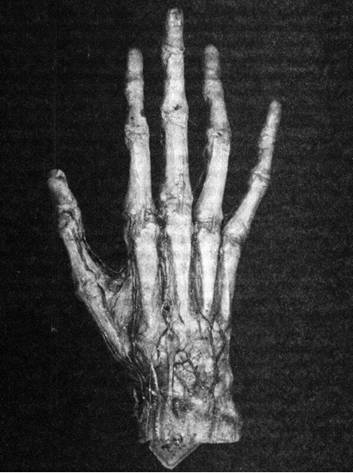
Илл. 65. Кисть руки человека
(Из выводов Г. Шеперда и И. Стрельникова не со всей полнотой явствует тот факт, что иная, чуть менее удачная конструкция передних конечностей навсегда оставила бы homo обычным животным, несмотря на всю развитость и «исключительность» его мозга.)
Вкратце рассмотрим анатомическую и нейрофизиологическую составляющие феномена кисти руки человека (илл. 65).
Эволюционный путь от эврибазальных складок древних рыб до кистей рук приматов понятен и в известной степени очевиден.
Примерно 450 миллионов лет назад эти складки начали то «разгоняться» в плавники, то редуцироваться, то переформировываться в жгутовидные гибкие лучи, присоски, лопасти, пальцевидные выросты и щупальца. При выходе части организмов на сушу плавники и лопасти перестроились вместе с плечевым поясом, частично погрузившись под кожу и образовав ключицы, частично выпятившись, чтобы обеспечить минимальную пальцеходность (илл. 66).
«Примитивные рептилии, в равной мере как и их непосредственные потомки — первичные млекопитающие, будучи ещё наземными формами и передвигаясь по зыбкой и неровной поверхности грунта, приобрели основные морфо-функциональные предпосылки, определившиеся приспособлением поверхности ладони к неровностям почвы. Это выразилось в пронаторно-супинаторной подвижности предплечья и хватательных свойствах кисти» (Jones W. Man's Place among the Mammals, 1929).
Co временем кости запястья и предплюсны удлинились, а сами конечности специфицировались в лазательные, хватательные, пальцеходные, роющие, стопоходящие и, соответственно, снабдились когтями, пальцами, копытами или волярными подушечками (илл. 67).
(Причины и механизмы этих изменений превосходно изложены в теории конечностей А. Н.Северцова, в его труде «Морфологические закономерности эволюции» (1939) и пересказывать их нет необходимости.)
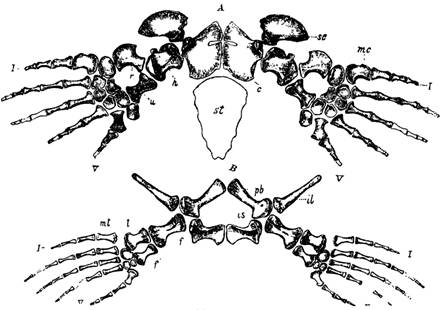
Илл. 66.
А. Плечевой пояс Edestosaurus dispar, вид сверху, с — coracoideum, sc — scapula, st — sternum, h — humerus, r — radius, u — ulna, me — metacarpus, l-V — пальцы.
В. Тазовый пояс Platecarpus simus, верхний мел.
Il — ileum, pb — pubis, is — ischium, f—femur, t — tibia, f — fibula, mt — metatarsus, l-V —пальцы.
Следует отметить, что с момента выхода на сушу, манипуляторная функция передних конечностей почти всегда «подразумевалась», но оставалась «бедным родственником» при простой локомоции и специальных функциях (вроде копания или присасывания к гладким вертикалям).
Исключение, как мы помним, составляли анатомические особенности примитивных архозавров, терапод, части зауроподоморфов, птицетазовых динозавров и рауизухий. (Именно на этих группах эволюция ставила свои «первые опыты» бипедальности и относительной свободы передних конечностей.)
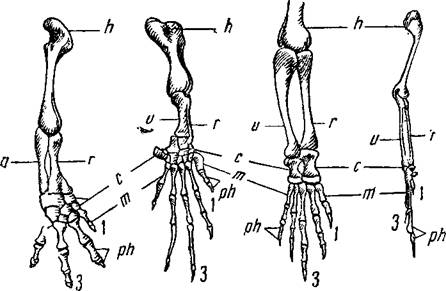
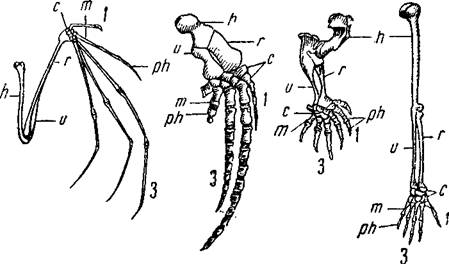
Илл. 67. Скелет передней конечности наземных позвоночных.
В верхнем ряду слева направо: саламандра, морская черепаха, крокодил,
птица. В нижнем ряду: летучая мышь, кит, крот, человек.
Гомологичные кости обозначены одинаковыми буквами.
К примеру, позднетриасовый Coelophysis. «Ключица сохраняется, но редуцирована, что высвобождает плечевой пояс и придаёт большую маневренность передним конечностям. Сами они слабые и данных об их участии в поддержании тела нет. Кисть, редуцированная до трёх функционирующих пальцев и рудимента четвёртого хорошо приспособлена для хватания» (Кэррол Р. Палеонтология и эволюция позвоночных, 1993). В. Кумбз полагает, что «пропорции конечностей некоторых динозавров очень близки к таковым у бегающих млекопитающих» (Coombs W. Theoretical Aspects of Cursorial Adaptations in Dinosaurs, 1978). Впрочем, всё не так однозначно: по большей части бипедальность динозавров отнюдь не означала приспособленность их передних конечностей даже к минимально сложным манипуляциям: к примеру, у мегалозавров передние конечности были трёхпалыми, снабжёнными длинными, загнутыми назад когтями, у тираннозавров — двухпалыми, а у части гадрозавров пальцы кисти были обвязаны перепонками. Осмотр черепа типичного терапода Allosaurus не предполагает возможности размещения в нём мозга, имеющего развитые зоны тонкой моторики (илл. 68). Мы очень мало знаем о мозге архозавров, терапод, зауроподоморфов и так далее, но сегодняшнее понимание того, как обширно должно быть церебральное обеспечение тонких манипуляций, позволяет (сопоставительно с черепами динозавров) усомниться в том, что они были способны на какие-либо движения, кроме самых примитивных и грубых.
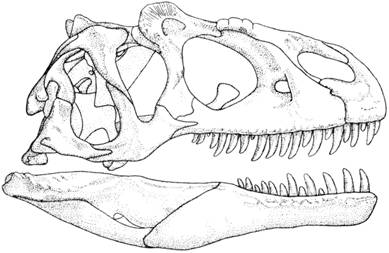
Илл. 68. Череп типичного терапода Allosaurus
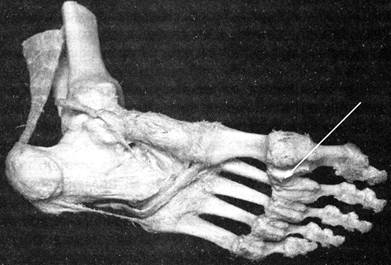
Илл. 69. Глубокая поперечная плюсневая связка —
lig. metatarsale transversum profundum
Ното стал «второй попыткой» эволюции полностью освободить передние конечности от опорных и двигательных нагрузок, но этого освобождения бы не произошло, и рука человека, несмотря на весь свой анатомический потенциал, никогда бы не состоялась как успешный манипулятор, если бы в стопе homo отсутствовала или не смогла бы развиться глубокая поперечная плюсневая связка — lig. metatarsale transversum profundum (илл. 69).
Эта связка, являясь фиброзным тяжем, идёт от головки первой плюсневой кости до пятой плюсневой кости, срастаясь с капсулами плюсне-фаланговых суставов, и связует собой головки всех плюсневых костей, формируя из них упругий свод. Если бы не этот свод, дающий упор не на всю подошвенную поверхность, а лишь на пяточный бугор и головки I и V плюсневых костей, продолжительное прямохождение было бы либо малореально, либо мучительно. Более того, именно эта связка, вкупе с подошвенным перекрестом hiasma plantare, апоневрозом и латеральным подошвенным возвышением eminentiaplantaris lateralis даёт стопе тонкую силовую маневренность (устойчивость) и преобразовывает энергию мышц самой стопы, голени и бедер в очень дозированные движения.
К примеру, анатомическая возможность обхватить горящую ветку пальцами руки, поднять её и начать с ней манипуляции — сама по себе мало что значит. Естественно, роль кисти руки в этом вопросе сверхважна, но только lig. metatarsale transversum profundum может обеспечить телу схватившего ветку то положение, когда огонь на ней не будет опасен для самого схватившего, то есть устойчивую и комфортную эректильность. Поясняю: обезьяна, проделавшая такой трюк с горящей веткой (по всей вероятности), подожжёт саму себя при первой попытке передвижения вперед. Впрочем, и настоящего прочного и легкого захвата ветви у обезьяны тоже не получится, но об этом — чуть позже.
Итак, как мы видим, полноценная манипулятивность передних конечностей невозможна без набора определённых анатомических свойств конечностей задних. В человеке эти два морфологических свойства «встретились» и начали очень любопытное взаимодействие. Полностью освобожденные руки позволили животному homo начать реализовывать на практике базовые возможности разума.
Переоценивать этот выигрыш homo в анатомической эволюционной лотерее, полагаю, не следует. Очень многие животные обладают удивительными качествами именно благодаря тому, что в их организмах «совпали» два, три или даже более взаимоработающие свойства. (Свойства могут быть малозначащими сами по себе, но в сочетании друг с другом — весьма продуктивными и сообщающими организму выживаемость и плодовитость.)
Вероятно, имеет смысл уточнить отличия кисти руки человека и обезьяны: прежде всего это (по Е. Даниловой «Эволюция руки» (1965)) удлинённость II пястной кости и редуцированность шиловидного отростка локтевой кости, что обеспечивает свободу кисти в ульнарном направлении и даёт исключительную свободу и мощь большому пальцу, мускулатура которого иннервируется сразу тремя (лучевым, срединным и локтевым) нервами.
Теперь вспомним основные задачи, которые решаются организмами через манипулирования. Это (по Шеперду) «схватывание, защипывание, разрывание, удерживание, выдавливание содержимого, раздавливание, обследование, ощупывание».
Нет необходимости уточнять, что смысл большинства этих действий — получение доступа к еде; посему основными манипуляторами становятся именно передние конечности как наиболее приближенные к ротовому отверстию.
Манипуляторы должны обладать не только значительной, но и тонко регулируемой мышечной силой, «обострённой» термической, болевой и тактильной чувствительностью, а проприоцептивная картина, транслирующая в мозг их положения, состояния, движения, — должна быть сверхточной55.
Всё это требует очень «подробной» иннервации их связок, мускулов и покровных слоев, установления мощных эфферентно-афферентных связей с головным мозгом и развития в самом мозге обширных корковых полей для обеспечения всех этих регулировок; а как мы знаем, увеличение головного мозга человека «шло не равномерно, а в основном за счёт увеличения мозжечка, двигательной коры и её ассоциативных областей» (Passingham R. Changes in the Size and Organization of the Brain in Man and His Ancestors, 1975).
Неслучайно в коре головного мозга человека именно кисть руки имеет наибольшее «представительство», во много раз превосходящее те моторные «зоны», что ведают стопой, бедром, голенью и так далее: «Следует отметить, что площадь соматотропических проекционных зон в коре полушарий пропорциональна сложности движений, выполняемых группой мышц. Наибольшую по площади соматотропическую проекцию имеют мышцы кисти» (Гайворонский И., Гайворонский Д. Функциональная анатомия нервной системы, 2007), а связь мозга и кисти обеспечивается (в том числе) частью пирамидного кортикоспинального тракта, состоящего из (примерно) миллиона волокон.
По Лассеку и Уитли (1945) — 1 100998 волокон; по материалам 23 исследований Ферхарта и Крамера (1952) — от 700000 до 1 282000 волокон; по Майеру (1959) — от 749000 до 1 391 000 волокон. Следует оговорить, что тонкая моторика рук — далеко не единственная «забота» tractus corticospinalis, а точное количество волокон, обслуживающих связь кистей рук с моторной корой, не установлено, но по мнению В. М. Бехтерева, подтверждённому исследованиями Д. Капперса, Г. Губера, Е. Кросби (1936), А.Лассека (1946), оно, вероятно, значительно. Примечательно, что пирамидный тракт человека, служащий, по мнению В. М. Бехтерева, для обеспечения точности и ювелирности «обособленных» движений (в том числе) пальцев, занимает до 30% площади поперечного сечения спинного мозга (по С. Блинкову, 1964). Сам В. М. Бехтерев пояснял столь значительное его представительство в спинномозговом субстрате следующим образом: «Пирамидный пучок является естественным последствием дифференцировки мозговых проводников, предназначенных для особой специальной цели, состоящей в передаче от коры к низшим центрам импульсов для обособленных движений. Поэтому вместе с большим развитием у животных обособленных движений всегда обнаруживается и большее развитие пирамидных пучков» (Бехтерев В. М. Основы учения о функциях мозга, 1905).
А здесь, полагаю, будет уместным возвращение к теме «белой крысы Олдса». Вернее, к той потребности головного мозга в раздражителях, что стала одной из вероятных причин филогенеза в целом и развития человека в частности.
Итак.
По всей вероятности, те возможности, которые дали животному homo первые манипуляции, т.е. способность пусть и минимально, но корректировать реальность, подчинять её, видоизменять предметы и вступать с ними в новые отношения, оказались ещё одним, новым, раздражителем, достаточно сильным для того, чтобы мозг человека начал искать этот раздражитель вновь и вновь, с той же настойчивостью, с какой мозг белой крысы в лаборатории Хэбба искал заветную педальку.
Возможно, в этом утверждении содержится излишний «физиологический механицизм», сводящий всю историю человека к поиску его мозгом тех раздражителей, что могли бы вызвать возбуждение нейронных масс и структур. Возможно.
И. П. Павлов в 1933 году характеризовал подобный «механицизм» следующим образом: «Многих, даже научно думающих людей почти раздражают эти попытки физиологического объяснения психических явлений, и поэтому эти объяснения сердито обзываются “механическими” с расчётом этим резко подчеркнуть как явную несообразность, нелепость сближения субъективных переживаний с механикой....Истинное механическое толкование остается идеалом естественно-научного исследования, к которому лишь медленно приближается и будет долго приближаться изучение всей действительности, включая в неё и нас. Всё современное естествознание в целом есть только длинная цепь этапных приближений к механическому объяснению, объединенных на всём их протяжении верховным принципом причинности, детерминизма: нет действия без причины» (Павлов И. П. Последние сообщения по физиологии и патологии высшей нервной деятельности, 1933. Вып. II).
Следует принять во внимание, что кроме предложенного нами «физиологического механицизма» ни одна из существующих гипотез антропогенеза пока не даёт ответа на очень важный вопрос: чем стимулировалось самое начало процесса камнеобработки, те самые первые опыты кидания, расколов и расщепов камней? (По всей вероятности, это «начало» было растянуто на десятки или даже сотни тысячелетий.)
Возможно, именно этот момент является «ключиковым» и ко всей дальнейшей истории человека, так как несколько проясняет сам принцип взаимодействия разума и анатомических особенностей homo.
Что именно могло побуждать обычное животное к его первым странным занятиям с камнями? Никаких объяснимых практических причин или стимулов делать это у него, по всей вероятности, не было. Тот бытовой результат, который мог бы прояснить заинтересованность homo в острогранных кварцах и гальках, мог образоваться и быть «обнаруженным» человеком палеолита лишь в результате множества проб и ошибок, спустя значительное время после начала его «экспериментов».
(Прогностические же способности чистого животного разума человека, как мы знаем, не велики, а при отсутствии в его гиппокампальной базе материала для конструирования ассоциаций — и вовсе ничтожны.)
Иными словами, животное homo не могло иметь даже малейшего понятия не только о конечном результате своих манипуляций с камнями, но даже и о результате промежуточном, так как не имело на тот момент опыта применения ни скребков, ни рубил. (Опыт придёт только с окончательным овеществлением и бытовым применением этих орудий, т.е. значительно позже.)
Попробуем восстановить эту картину и найти наиболее возможный стимул.
Итак, ряд особенностей кисти передней конечности когда-то дал животному homo возможность прочно сжать камень, а затем и кинуть его. Будучи целым комплексом необычных зрительных, миологических и тактильных впечатлений, это действие должно было стать очень сильным новым раздражителем; вероятно, прежде всего за счёт вдруг возникшей «баллистической власти» над камнем, того острого и странного ощущения, когда уже отделившийся (ушедший в полет) предмет продолжает «исполнять желание» метнувшего его, двигаясь по заданной траектории.
Удивительность ощущения этой власти над предметом (новизна и сила раздражителя), несомненно, спровоцировала мозг на многократные повторения этого действия.
А потом настало мгновение, когда брошенный камень ударился о другой камень и раскололся.
Пришла пора подобрать осколки и убедиться в инструментальных возможностях разбитого камня, теперь имеющего острую грань, что крайне важно для существа без настоящих клыков и когтей.
Эти принципиально новые для животного действия спровоцировали возникновение целого ряда новых сильных раздражений его мозга. Многократное усугубление возможностей, которое дали осколки, «отворило врата» в целый мир необыкновенных по силе возбудителей, за которыми и начал «охоту» мозг.
Несомненно, опыт метания и разбития был многократно повторен. Потом пришло видение того, что метательный способ часто приводит к потерям ценных осколков. (Разум животного здесь выполнил вполне посильную для него работу, так как все компоненты задачи были явными.)