Неслучайно все подобные теории (и Э. Кэндела, и подобные им) тщательно «обходят» или игнорируют вопросы функциональности трактов, кортикофугальные и кортикопетальные связи, общую активирующую силу ретикулярной формации, сложные роли таламуса и гиппокампа, да и вообще всю эволюционную архитектуру мозга.
Данная ремарка о «синапсизме» ни в коем случае не ставит под сомнение важность и изящество исследований Кэндела о расширении синапса, соединяющего сенсорные и двигательные нейроны. Его эксперименты с аплизией (Aplysia) а впечатляют, но их трактовка, возможно, является несколько поспешной, а в силу этого — не вполне точной. Впрочем, как мы помним, в истории исследования мозга это не первый пример такой поспешности, посему основанием для обобщающих гипотез всё же целесообразнее брать не «модные» версии, а фундаментальную классику. Нобелевская премия 2000 года, которой был удостоен за свои исследования Э. Кэндел, несомненно, наивысшим образом сертифицирует его исследования, но стоит помнить и о премии 1949 года, полученной Антониу Каэтану ди Абреу Фрейри Эгаш Мониш (лоботомия).
а Морская улитка. — Прим. ред.
Вернёмся к нашей теме о физиологически парадоксальной потребности мозга в максимально активирующих его факторах. Это позволит аккуратно и последовательно перейти к рассмотрению вопросов «разума» и «интеллекта».
Из всех существующих ныне теорий ни одна не способна предложить объяснение «парадоксу активации», но вот версия У. Г. Пенфилда — Г. Джаспера может отчасти её проиллюстрировать. Это немного, но это всё, чем мы располагаем на данное время. (Если не опускаться до фантазий.)
К слову, корректность такого наименования данной гипотезы сомнительна. Её многие фундаментальные положения базируются на исследованиях С. Кобба (1944), Г. Бертранда (1956), Г. Мэгуна (1950,1952, 1954), и лишь отчётливость она приобрела в книге У. Пенфилда и Л. Робертса «Речь и мозговые механизмы» (1959).
Версия не бесспорна, но предельно проста и с честью проходит апробацию логикой эволюционности.
По У. Пенфилду — Г. Джасперу все новейшие корковые образования есть естественная клеточная «проекция» различных зон ствола мозга, которые тот «разращивал» в процессе эволюции как собственный тонкий инструментарий.
Более того (согласно схеме, илл. 61), каждое функциональное поле коры образует единицу со стволовыми структурами и рассматривается как целое. «Таким образом в коре находятся четыре больших поля, осуществляющие то, что может быть названо главной линией связи с окружающим миром» (Cobb S. Fundamentals of Neuropsychiatry, 1944).
Происхождение этих четырёх «единиц» увязано с развитием основных рецепторов, с усложнением и многообразием воспринимаемых ими раздражений и, соответственно, с потребностью в развитии анализаторов: «Большие поля коры мозга человека, находящиеся в височной, передней лобной и задней теменной областях, соединены в мозгу между собой, что делает возможным выполнение функций, которые скорее могут быть описаны как психические, чем моторные и сенсорные. Все поля коры объединяются с субкортикальным серым веществом при помощи специфических и неспецифических нервных волокон проекционных систем» (Джаспер Г. Подкорковые взаимосвязи, 1954).
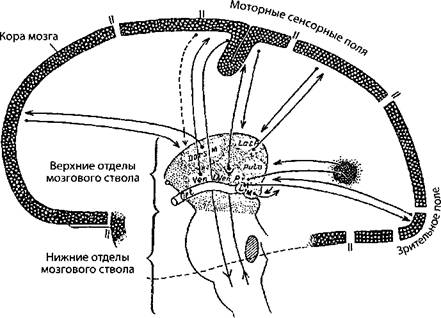
Илл. 61. Диаграмма иллюстрирует гипотезу, согласно которой каждое функциональное поле коры образует единицу с некоторыми частями диэнцефалона, проекциями которых она становится в процессе развития (по Пенфилду)
Меж схемой У. Пенфилда — Г. Джаспера и «парасагиттальными» схемами мозга крысы Дж. Олдса трудно не увидеть косвенную, но любопытную связь. Обе схемы, даже если и не являются «ключиками» друг к другу, то по крайней мере друг друга иллюстрируют.
Рассмотрим их.
На всех схемах (см. илл. 60 и илл. 61) мы видим примерно одни и те же области: таламус, гипоталамус, гиппокамп, мамиллярные тела, пульвинар, межножковую ямку, ядра ретикулярной формации и так далее.
Как показывают опыты Дж. Олдса и его последователей, именно эти древние участки проявляют «жадность» к раздражениям, а получив их, отвечают чрезвычайно мощной, концентрированной рефлекторикой, резко меняющей обычное состояние мозга.
На схеме У. Пенфилда — Г. Джаспера те же самые анатомические участки мозга заявлены как локализаторы генерализующей нейронной «силы», которая и «выстроила» вокруг себя полушария мозга, снабжённые самыми различными проекционными, ассоциативными, тонкомоторными и пр. центрами.
Эту мысль У. Пенфилда — Г. Джаспера подтверждает классика цитоархитектонических исследований, зафиксировавшая плавную, но отчётливую и явную этапность развития мозга: «Как показывают наши исследования, в области древней, старой и межуточной коры формации переходят одна в другую путём последовательных переходов и в то же время прерывисто, так как и формации, представляющие собой эти переходы, отделены друг от друга линейными и резкими границами» (Филимонов И. Н. Цитоархитектоника. Общие понятия. Классификация архитектонических формаций // Цитоархитектоника коры большого мозга человека, 1949).
Итак.
На основании опытов Дж. Олдса и его последователей мы с известной долей уверенности можем предположить наличие в диффузных нейронных группах и ядрах мозгового ствола потенциал поиска интенсивного раздражения. Учитывая сопоставимость конструкции ствола по принципиальным признакам у всех позвоночных (от самых примитивных до высших млекопитающих), допустимым будет и предположение, что это свойство характерно для древнейших церебральных структур на всей протяженности истории головного мозга, начиная с протерозоя.
Что могло бы служить этим раздражителем?
В первую очередь это внешняя информация, являющаяся единственным и вечным «партнёром» нервной системы и превосходным глобальным раздражителем.
К слову, под «внешней» я подразумеваю любую среду, находящуюся за пределами самой нервной системы, т.е. и иннервируемые ею органы и ткани.
Лучшее подтверждение тому — история рецепций, начавшаяся в протерозое. На первоначальном этапе, когда нервная система была диффузна и слаба, ей вполне хватало тех несильных раздражений, которые предлагали термо-, хемо- и механорецепции.
По мере её укрупнения и формирования зачатков протоцеребрума — мы всё отчётливее видим растущий «аппетит» усиливающихся нейронных скоплений к раздражениям.
Этот «аппетит» спровоцировал появление таксисов, кинезисов, дорсальную световую реакцию, диффузную светочувствительность и, наконец, образование фоторецепторов самого разного типа, от шестого брюшного ганглия речного рака до глаза ястреба (Falco).
(То, что справедливо в отношении света как превосходного и многоликого раздражителя, может быть отнесено и ко всем остальным рецепциям, имеющим примерно такую же историю.)
Логично предположить, что чем крупнее и совершеннее становилась нервная система, тем более сильное раздражение ей требовалось. Яркость, точность, многообразие внешней информации, поставляемой рецепторами, вероятно, обеспечивали его силу и непрерывность. Это раздражение провоцировало рост, а рост — требовал всё более сильного раздражения.
Вероятно, в истории мозга это была самая прекрасная и фантастическая эпоха. Грей Уолтер, в своё время отметив сходство электрических разрядов, регистрируемых в нервной цепи медузы, с разрядами, возникающими у человека во время эпилептического припадка, очень поэтично резюмировал результаты своих наблюдений: «Возможно, при эпилепсии мы наблюдаем оставшийся от далеких времён отголосок этого первого нервного урагана» (Уолтер Г. Живой мозг, 1966).
В пермском периоде, как следствие этого всё возраставшего «аппетита» сформировавшихся нейронных масс к раздражению, вероятно, и началась кортикация (усовершенствование рецепторов и развитие проекционных и ассоциативных зон), по мере которой картины свирепого мира палеозоя, а затем и мезозоя — становились раздражителем всё более сильным и многообразным.
Возможность взаимодействия организма с реалиями среды (предметами, явлениями, свойствами, катаклизмами) оказалась ещё одним мощным раздражителем. Вероятно, это сдетонировало нарастание тонкомоторных зон коры головного мозга. Развитие этих зон, обогатив поведение, обеспечило большую событийность в жизни организма, что тоже стало сильным раздражителем и спровоцировало организацию ещё более тонких потенциалов коры.
Это положение имеет подтверждение в работах D. Ferier (1876), Р. Flechsig (1876), К. Brodman (1909), Н. Stephanet O. Andy (1964), R. Lorente de No (1947), G. Bishop (1961), J. Papez (1961), В. M. Бехтерева (1906), И. H. Филимонова (1949, 1963), Г. Полякова (1964), С. Саркисова (1964), а наиболее чётко было сформулировано в труде А. И. Карамяна «Эволюция конечного мозга позвоночных» (1976):
I. «...Были установлены факты, свидетельствующие о том, что в восходящем ряду млекопитающих, наряду с увеличением площади новой коры отмечается тенденция к более узкой и чёткой локализации моторных, сенсорных, зрительных и слуховых функций».
II. «В процессе фило- и онтогенической эволюции коры головного мозга отмечаются возрастающее усовершенствование её ассоциативных и проекционных полей, дифференцировка нейронной организации...».
Так что, вполне возможно, что белая крыса Олдса своим «загадочным» поведением в 1954 году намекнула нам на то, что история мозга — это в первую очередь поиск и нахождение раздражителей. (Что, впрочем, вполне увязывается как с «конфликтной специализацией» нейрона, так и со зримой «этапностью» церебральной архитектуры и «проекционной» теорией У. Пенфилда — Г. Джаспера.)
Попутно следует отметить, что «пенфилдовская» теория «клеточной проекции» способна ответить на один из самых щекотливых вопросов нейрофизиологии — о причинах отсутствия в коре мозга центра (или центров) локализации или хотя бы анализатора такого сверхважного фактора, как «боль».
Мы уже вскользь упоминали об этом факте, основываясь на мнении И. С. Бериташвили: «Боль не имеет своего представительства в новой коре больших полушарий» (Бериташвили И. С. Структура и функции коры большого мозга, 1969).
К мнению Ивана Соломоновича можно добавить и высказывание Ч. С. Шеррингтона: «Ни одна из областей коры не может считаться областью, где локализуется ощущение боли» (Шеррингтон Ч. С. Интегративная деятельность нервной системы, 1969), а также весьма авторитетные версии В. Кэннона (иссл. 1914,1922,1928 гг.), С. Хоскинза (1912), П. Анохина (1956), С. Окса (1965), С. Серебренникова (1932), А. Данилова (1934), Д. Гзгяна (1938) и отчасти Л. О. Орбели (1938).
Общий опыт исследования боли в лаборатории Л. О. Орбели был в свое время обобщен М. Панкратовым (1934), доказавшим, что не только декортикация, но и полное удаление полушарий не снимает болевой чувствительности.
Что такое боль?
Грубо говоря, это генеральный физиологический индикатор, подсказывающий организму приближение к увечью или смерти или удаление от них; это важнейшая характеристика многих явлений, предметов и обстоятельств.
Боль руководит не только поведением организма в целом, но и самой рефлекторикой (т.е. поведением целых групп нейронов, секрецией, инкрецией, мобилизацией адренергических механизмов и т.д.).
В организме трудно найти более «авторитетный голос», чем «голос боли».
Её механизм достаточно хорошо изучен: «Импульсы, возникающие при болевых раздражениях, направляются прежде всего в зрительный бугор и в гипоталамическую область, а оттуда в ретикулярную формацию, благодаря чему возникает генерализованное влияние на всю мозговую кору» (Анохин П. О физиологических механизмах болевых реакций, 1956).
Мнение П. Анохина (в общем и целом) подтвердилось в исследованиях Г. Френча, М. Верзано, Г. Мэгуна (1953), А Ардуини и М. Ардуини (1954), Кагена (1955), Г. Мэссона (1956), Кила (1957) et cetera.
Сидней Окс в своих «Основах нейрофизиологии» (1969) дипломатично обобщил все имеющиеся точки зрения: «Можно предположить, что восприятие боли происходит на подкорковом уровне».
С. Н. Оленев (1995) и G. Battaglia (1992) предложили ещё более тонкую и конкретизированную схему её локализации — это каудодорсальный отдел латеродорсального сектора substantia grisea centralis, расположенный вокруг водопровода среднего мозга и интраламинарных ядер таламуса.
Возможно, точки локализации боли остаются (отчасти) дискутивным вопросом, но уже понятно, что все они располагаются исключительно в древних, преимущественно таламических структурах и ядрах ретикулярной формации. (Там же, по всей видимости, происходят и основные адренергические реакции, связанные с болью.)
Иное (исходя из эволюционной логики) трудно было и предположить; боль является настолько важным регулятором поведения, что представить себе развитие жизни без её участия категорически невозможно.
При всей кажущейся непререкаемости этого явления боль не следует абсолютизировать; во многих случаях ведущие агрессии легко вступают с ней в конфликт и «берут на себя» управление поведением. Но и при этом боль сохраняет важность роли, выступая уже в качестве усилителя агрессии.
Естественно, как явление, боль — ровесница самых первых, докембрийских ганглиозных начатков мозга, а окончательное развитие и оформление генерирующего её субстрата приходится на последующие 300 миллионов лет, т.е. на «докортикальный» период.
Именно по этой причине кора, являющаяся «клеточной проекцией» (инструментом) стволовых структур, которая появляется только в пермском периоде у терапсид и пеликозавров и не имеет ни центров, ни анализаторов боли, как, впрочем, и многих других сверхважных функций; все они обрели свой материальный субстрат до зарождения и развития коры.
Напомню, что помимо боли, вне прямой «компетенции» коры находятся агрессии, память, сознание, сон, бодрствование, функция «личности», практически все висцеральные регуляторы, грубая моторика и, по всей видимости, т.н. разум, т.е. все те качества, которыми, без сомнения, должны были обладать существа «докортикального» периода.
На чём может быть основана наша уверенность в наличии вышеперечисленных свойств у тех, кто населял землю в кембрийский, ордовикский, силурийский, девонский и каменноугольный периоды (а это в сумме 250 млн лет)?
Эта уверенность может быть основана на множестве фактов, самый доступный из которых — отражение вашего собственного лица в зеркале.
Вглядитесь.
Вы увидите там любопытный эволюционный продукт, несущий на себе «печати» всех этапов филогенеза за 500 миллионов лет.
К примеру:
Амфибии девона оставили на этом лице т.н. мигательную глазную перепонку — полулунную складку (plica semilunaris conjunctivae) во внутреннем углу глаза.
Жаберная дуга костных рыб силурийского периода определила форму и особенности нижней челюсти этого лица. (Как, впрочем, и у всех поперечноротых.)
В конструкцию же самого глаза хаотично «вложились» эпохи от раннего кембрия до мела, обеспечив наш глаз набором конструкционных нелепостей, вроде вывернутой наизнанку сетчатки, слепого пятна и проходящих сквозь сетчатку множественных сосудиков, существенно темнящих и искажающих получаемое изображение.
Не вполне академично, но тем не менее точно охарактеризовал странности глаза человека Т. Лэм (Австралийский нац. унив., Центр по совершенствованию наук о зрении): «Фактически, если бы глаз со всеми его недостатками спроектировали инженеры, они были бы уволены» (Лэм Т. Эволюция глаза //ВМН, 2012).
Напомню, что когда наш объект рассмотрения ещё был «личиком» хвостатого четырехмесячного эмбриона, то волоски на нём располагались группками по «три» и по «пять», что в точности соответствует расположению чешуек у рептилий каменноугольного периода, а ещё чуть раньше кожа на нём была совершенно прозрачна, так как состояла из двух слоев клеток (кубических и плоских), как у низших жаберных позвоночных ордовика (илл. 62).
Таких «печатей», повторяю, множество, наличие их давно является общим местом, но мы вспомнили о них только в контексте поиска доказательств наличия сознания, памяти, «разума», «личности» у тех существ, что населяли землю в докортикальный период.
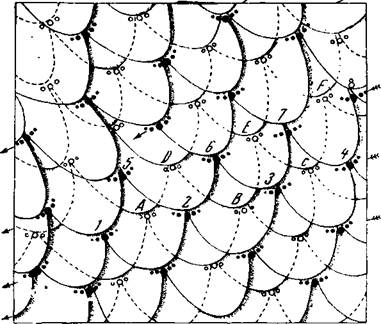
Илл. 62. Схема расположения зачатков волос на коже туловища человеческого зародыша: реконструкция чешуевидного характера поверхности кожного покрова четырехмесячного человеческого плода. Видны группы по три и пять волосков (по Штёру)
Каким образом может быть взаимоувязано проявление предковых черт у homo и наличие сознания, памяти и разума у существ силура и ордовика?
Самым прямым.
Полагаю, рудименты и атавизмы — это прежде всего напоминание о биологической успешности тех видов, родов и семейств, что были сильны и долгожительны настолько, что смогли транслировать свои черты в организмы даже самых далеких потомков.
А сила, успешность, долгожительность вида (рода или семейства) — это прежде всего свидетельство его способности выстроить точное поведение в ответ на все «вызовы» среды и практиковать его в течение нескольких миллионов лет, через благополучие вида, рода или семейства не просто закрепляя свои морфологические особенности, но и делая их «неизбежными» для всей последующей эволюционной цепочки.
Излишне говорить, что такое поведение невозможно без точной и адекватной оценки реальности, которая, в свою очередь, невозможна при отсутствии памяти, полноценного сознания и того, что называется «разумом».
Мы уже говорили об «обязательности» сознания для всякого живого существа и о том, что качество этого сознания зависит исключительно от развитости рецепторов и проекционных зон (глава VIII).
Нет никаких оснований предполагать, что для организмов палеозоя этот принцип недействителен.
Более того, при желании мы можем обратиться к классике и получить весомые подтверждения справедливости его применения, основанные на долгой и ответственной экспериментальной работе с низшими позвоночными. Рыбы: А. Карамян (1956, 1970), Ю. Фролов (1926), Н. Сихарулидзе (1967), Ю. Холодов (1963); амфибии: И. Бериташвили (1929), К. Леутский (1929), Н. Бебуришвили, Н. Чичинадзе (1936), В. Бианчи (1967); рептилии: К. Поляков (1930), Н. Wagner (1933), Е. Diebschlag (1934), А. Эльдаров (1968). Полагаю, что наиболее точными и лаконичными являются выводы И. С. Бериташвили: «У низших позвоночных сенсорные элементы в головном мозгу, несомненно, имеются. Они производят ощущения внешнего мира, воспринятого через рецепторы. У них также имеются интегрирующие нейроны, которые объединяют эти сенсорные элементы и тем самым производят восприятие конкретных объектов. При этом мы все хорошо знаем, что эти животные — рыбы, амфибии, рептилии и птицы — не только воспринимают внешние объекты, но и правильно проецируют их во внешней среде, и это даёт им возможность развивать адекватную двигательную активность в отношении воспринятых объектов...» (Бериташвили И. С. Память позвоночных животных, её характеристика и происхождение, 1974).
Теперь рассмотрим память.
Когда мы упоминаем о ней применительно к организмам кембрия, ордовика, силура, девона и карбона, то не возникает особых противоречий с каноническими представлениями об эволюционной последовательности цереброгенеза и об изначальном распределении функций в структурах мозга.
Данное утверждение легко иллюстрировать стадиями развития гиппокампа, остающегося и по сей день основным обеспечителем такого важного фактора, как память.
Зачатки гиппокампальных структур определяются уже у миксин (мощными слоями пирамидных клеток) и становятся вполне зримыми у челюстных поперечноротых; а открыв мозг амфибии сперва на уровне прекомиссуральной области переднего мозга, а затем на уровне интервентрикулярных форамин, мы отчётливо увидим и пирамидки primordium hippocampi, и кривые пластиночки fibrae hippocampi extemae.
У рептилий картина анатомического оформления гиппокампальных структур становится ещё ярче. Её превосходно описал Е. Сепп: «Область, которая у лягушки была представлена малодифференцированным primordium hippocampi, у ящерицы дифференцирована на слой пирамид аммонова рога и зернистый слой — fascia dentata» (Сепп Е. История развития нервной системы позвоночных, 1959).
Впрочем, наличие образной памяти у амфибий и рептилий никогда и не являлось дискутивным вопросом.
И. С. Бериташвили, основываясь на собственном обширном экспериментальном материале, пишет: «Образная память у амфибий, как выяснилось, развита в значительной степени, но хвостатые амфибии (тритоны) отстают от бесхвостых (жабы и лягушки), а из бесхвостых жабы стоят на более высокой ступени, чем лягушки» (Бериташвили И. С. Память позвоночных животных, её характеристика и происхождение, 1974).
Рептилии, как существа более тонко и сложно организованные, чем амфибии, демонстрируют уже не только образную, но и эмоциональную память, причём частичная децеребрация подтверждает «гиппокампальную версию» её «обитания». «Итак, у ящериц без переднего мозга совершенно исчезала как образная память, так и эмоциональная память страха» (Бериташвили И. С. Память позвоночных животных, её характеристика и происхождение, 1974).
К слову напомню, что именно «у рептилий впервые в ряду позвоночных оказывается возможным образование временных связей типа ассоциации» (Карамян А. Функциональная эволюция мозга позвоночных, 1970).
Как вы помните, третья позиция, которую мы применили для характеристики организмов докортикальной эры, был т. н. «разум».
Кавычки, вероятно, можно снять, так как мы употребляем это понятие не в его литературном, а строго в нейрофизиологическом смысле, определяя с помощью данного термина способность мозга к совершению правильной оценки предложенных реальностью обстоятельств.
Мы уже говорили о разуме в главе X как об операции сопоставления данных сознания и данных памяти. Результатом этого молниеносного сопоставления является точность поведенческого акта. (Применительно к предмету нашего обсуждения больше подходит определение «чистый разум», несмотря на некоторую использованность этого словосочетания ещё в 1781 году.)
Иными словами, «чистый» разум — это не «роскошь» мозга, а весьма тривиальный продукт его самых древних механизмов; ведь точность поведения, которую способны обеспечить только оценочные возможности разума — это вопрос элементарного выживания, т.е. самым простым доказательством наличия разума служит факт сохранности и благополучия организма и, соответственно, вида (рода, семейства).
Вероятно, принцип «чистого» разума неизменен со времён раннего кембрия и является идентичным у всех организмов.
Естественно, за сотни миллионов лет по мере развития рецепторов совершенствуется сознание, а развитие гиппокампальных, лимбических, корковых формаций, скорость кортикопетальных и кортикофугальных связей расширяет базы памяти и ускоряет к ним доступ.
Совокупность этих обстоятельств, разумеется, развивает разум, который есть обязательный результат взаимодействия всех этих структур.
В данном контексте знаменитый «шокирующий тезис» И. П. Павлова, озвученный им на XIV международном физиологическом конгрессе (Рим, 1932), получает, вероятно, свое объяснение. Напомню: «В окончательном варианте большими полушариями собаки постоянно производится в разнообразнейших степенях как анализирование, так и синтезирование падающих на них раздражений, что можно и должно назвать элементарным, конкретным мышлением».
Здесь следует обратить внимание на следующие важные нюансы:
I. Данный постулат является не просто следствием тридцатилетней экспериментальной работы И. П. Павлова, но и тем итоговым выводом, что прямо противоречит его же теории «второй сигнальной системы у человека».
II. Этот тезис И. П. Павлова лишний раз указывает на важность соблюдения терминологической аккуратности. Примерно представляя механизмы «внутренней речи» (мышления) и её происхождение, гораздо точнее было бы употребить вместо термина «мышление» понятие «разум», как обозначитель естественного, а не искусственного, «запускаемого» только внешним специальным научением процесса. И. М. Сеченов не без гордости и достаточно категорично подчеркивал: «Мы — не философы», тем самым отказывая нейрофизиологии в праве на малейшую неточность или расплывчатость формулировок.
Возможно, на данное время данное понятие (разум) уместнее всего применять только к животным, так как у современного homo он настолько растворен в мышлении и интеллекте, что практически не поддаётся отпрепаровке от этих двух сложных и эффектных продуктов работы мозга. (Разумеется, он может быть отчётливо наблюдаем и у человека, но лишь в тех редких случаях, когда тот вырастает в полной изоляции от речи, ритуалов, игр, мифологии и прочей социальной атрибутики homo).
Тем не менее очень трудно избавиться от соблазна рассмотреть особенности разума на примере социализированного homo, учитывая, что именно данный вид является объектом настоящего исследования.
Впрочем, ничто и не мешает нам сделать такую попытку.
(Чистоту нашего логического эксперимента возможно обеспечить лишь очень приблизительно.)
Нам следует из традиционной картины представлений homo о себе и мире аккуратно удалить всё то, чему человек обязан современному интеллекту, с его обширными информационными накоплениями.
Для этого, впрочем, нет необходимости «возвращать» homo в палеолитическое (или мауглеоидное) состояние.
Достаточно взять в качестве образчика «чистого человеческого разума» мироощущение аборигена Новой Гвинеи, Австралии, рыболова Огненной Земли, охотника западного побережья Африки или крестьянина из России начала XX века.
Эти представители homo хороши тем, что очень подробно и авторитетно описаны классиками антропологии, в частности, скрупулёзно задокументированы их анатомо-физиологические воззрения, являющиеся, безусловно, ключевыми в деле познания и понимания мира.
Несколько удивляет схожесть этих воззрений у народов, «разнесенных» географически и до середины XX века не «соприкасавшихся» своими культурами.
Это удивление, впрочем, было сформулировано ещё в 1922 году Л. Леви-Брюлем: «По мере того, как исследователи обнаруживали, или, вернее, изучали народности низшего типа в самых отдалённых, а иногда совершенно противоположных пунктах земного шара, вскрывались поразительные аналогии между некоторыми из этих народностей, доходившие порой до полного сходства в мельчайших деталях» (Леви-Брюль Л. Первобытное мышление, 1930) и отчасти объяснено И. М. Сеченовым: «Основные черты мыслительной деятельности человека и его способности чувствовать остаются неизменными в различные эпохи его исторического существования, не завися в то же время ни от расы, ни от географического положения, ни от степени культуры» (Сеченов И.М. Избранные труды, 1935). Иван Михайлович, к сожалению, не дал подробной трактовки своему же постулату, и чуть ниже мы будем вынуждены сделать это сами.
Антропология объясняет эти сходства примерно одинаковой степенью религиозности, социализации и общего развития.
Нейрофизиология, вероятно, могла бы предложить иное, ещё более простое объяснение: образчики чистого разума представителей разных этносов столь схожи, поскольку являются продуктом идентичных физиологических процессов головного мозга, а вызывающие их факторы для всех народов примерно одинаковы, а набор их невелик и тривиален: жизнь, смерть, труд, война, голод, деньги, половые отношения, болезни, дети, имущество и так далее. (Климатические, этнографические и прочие различия, конечно, привносят некоторую вариативность, но она ничтожна.)
Попытки обнаружить структурное различие меж мозгом европейца и (к примеру) африканца, популярные во времена Грациоле, Лайеля, Оуэна, Маршалла (это середина XIX века), закончились закономерным конфузом. Отсутствие сколько-нибудь существенных размерных отличий меж среднестатистическим мозгом европейца, африканца, аборигенов Австралии, Восточной Азии и представителей арктических народов, было доказано С. Мортоном (1849), П. Тобиасом (1970), К. Смитом и К. Вейлсом (1990) и пр. (Morton S. Observations on the Size of the Brain in Various Races and Families of Man (1849); Tobias P. Brain Size, Grey matter and Race — Fact or Fiction? (1970); Smith C, Beals K. Cultural Correlates with Cranial Capacity (1990)).
Впрочем, эта ремарка — простая дань академизму: отсутствие какой-либо связи меж размером мозга и его возможностями обсуждалось в настоящем исследовании (глава II) и является не просто доказанным нейрофизиологическим фактом, но и общим местом. Напомню основные исследования, которыми мы руководствуемся в данном вопросе: С. Р. Кахаль (1911), И. Филимонов (1940), О. Адрианов (1993), М. Нестурх (1967, 1970), В. Эфроимсон (1964), Г. Рогинский (1948), A. Reidel, С. Harper, Н. Kretschmann (1994), М. Плисецкий (1956), Г. Левин (1963), К. Beals, С Smith, S. Dodd (1984), М. Hennenberg (1984), М. Lathan (1974), D. Halpern (2000), J. Graves (2002) et cetera.