Следует понимать, что вопрос «зачем» ответа на сегодняшний день не имеет; более того — никаких, даже самых кружных, дальних и туманных путей к нему не «просматривается».
Тем не менее, его не следует игнорировать, т.к. он косвенно, но очень внятно указует на ограниченность исследуемого нами явления, i.e. интеллекта homo, следовательно, некие свидетельства о природе и свойствах интеллекта всё же содержит. (Ad hinc ei recte maxime.???)
Классический эволюционизм, биология, палеонтология, палеозоология, геология, зоология, антропология, физика и химия в известной степени отвечают на вопрос «как развивалась жизнь».
И лишь нейрофизиология способна ответить на третий по степени важности вопрос, «почему» она развивалась, и что было её детонатором и стержнем.
Непосредственно само развитие жизни из аморфных клеточных субстанций, возможно, начинается только с развития нервной системы и ею же определяется, так как сама по себе активная жизнь (в известной степени) есть реакция клеток на воздействие среды. Нервная система из клеточного «сырья», как чеканщик из металла, формирует организмы, сложность которых усугубляется по мере развития самой нервной системы.
Здесь вновь будет уместно возвращение к теме «биологической индивидуальности», т. н. личности организма, ибо только её присутствие способно объяснить ряд важных моментов.
Тут мало уместно понятие «страстность»50, но русский язык небогат, и придётся употребить именно это нелепое слово, чтобы хоть как-то характеризовать ту побуждающую силу, что провоцировала зачаток нервной системы совершенствоваться и разрастаться.
Слепому, глухому, немому, лишённому обоняния организму эпохи докембрия — страстно потребовалось расширение своих возможностей, чтобы нормально убивать и размножаться, вовремя рождаться и вовремя умирать, исполняя неведомую эволюционную миссию.
Бесспорно то, что организм получил эти возможности только благодаря развитию нервной системы, усложнение которой — усложняло и развивало носящий её организм, провоцируя образование рецепторов, органов, органокомплексов, эндокринных, иммунных и иных механизмов. Разумеется, усложнение и развитие было бы невозможно без интегрирующего начала, способного контролировать и направлять развитие самой нервной системы, и неуклонно ведущего её к формированию сложного и всевластного органа, который мог бы оптимально управлять всем организмом. (Головного мозга.)
Стоит отметить, что развитие нервной системы даже и в масштабах эволюции, вероятно, всегда было «опережающим»; совершенствуясь и усиливаясь, именно нервная система «тянула» за собой организм, провоцируя его морфологические новации и перестройки.
В известной степени это утверждение (согласно теории рекапитуляции*) — иллюстрируется эмбриогенезом современного homo, в котором закладки всего, имеющего отношение к мозгу и НС, существенно «опережают» развитие всех прочих структур.
*Краткое и быстрое повторение, воспроизведение основных этапов развития предковых форм в ходе индивидуального — особенно зародышевого — развития у современных взрослых организмов.
Академическая нейрофизиология комментирует факт данного несомненного «опережения» следующим образом: «Развивавшийся в эволюции мозг сам становился фактором усиления и ускорения видообразования и эволюции» (Стрельников И., акад. Анатомо-физиологические основы видообразования позвоночных, 1970); «Мозг, его полушария — в известной степени орган для беспрерывного дальнейшего развития животного организма» (Павлов И. П. Избранные произведения, 1949).
И. П. Павлов и И. Стрельников говорят, впрочем, уже не о нервной системе, а о мозге. Сути вопроса это не изменяет, но в известной степени обязывает совершить краткий экскурс в историю формирования головного мозга.
Чуть выше мы определили примерное начало развития «биологической индивидуальности» границей эдиакарского и кембрийского периодов.
А теперь, «перепрыгнув» через следующие сто миллионов лет, кратко, но ясными словами очертим историю возникновения головного мозга позвоночных, в котором, собственно, интересующая нас функция, т.е. «биологическая индивидуальность», или «личность», и расположилась со всем возможным удобством.
(Опустим этап оформления спинного мозга в тяж, защищённый позвонками. Тут всё относительно понятно и даже отчасти уже являет собой общее место.)
Итак.
Всё «таинство» происхождения головного мозга — это простое «загущение» нервных волокон спинного мозга в одной точке.
В своё время (вероятно, в протерозое) это ничтожное тогда «загущение» сконцентрировалось именно в той части организма, которая чаще находилась «впереди» во время движения и, соответственно, чаще имела дело с препятствиями, едой, противником или объектом совокупления.
К слову, у вендобионтов (жителей эдиакарского «рая»), таких как Dickinsonia costata, Aspidella, Tribrachidium, по большей части ещё не существовало «переднего» или «заднего» конца (илл. 52).
Поначалу образующееся нейронное и рецепторное скопление давало лишь усиление тактильной чувствительности в «переднем» сегменте организма, но в слепом мире конца протерозоя — начала кембрия даже это обеспечивало значительное преимущество.
Процесс развивался, на волне успеха концентрация усугублялась, провоцируя постепенное образование принципиально новой анатомической формы.
Нет сомнения, что первыми её границами были три оболочки спинного мозга, а сама форма была предельно скромна. (По крайней мере, значительно скромнее, чем то, что мы видим уже в среднем палеозое, если судить по неизменному с тех пор мозгу целаканта Coelacanth).

Илл. 51. Схематическое изображение гипотетического протокраниата (по Северцову)

Илл. 52.Tribrachidium. У этого вендобионта хорошо видно отсутствие «переднего» и «заднего» конца
Вероятно, это было лишь некое ничтожное утолщение, масштабированно повторяющее геометрию спинного мозга, со всеми его фиссурами и бороздами — передней, задней и латеральными.
Трудно сказать, сколько именно миллионов лет потребовалось на то, чтобы этому «сгусточку» стало бы тесно в рамках простого утолщения рострального конца спинного мозга. (Вероятно, около 100 млн. лет, если считать от кембрия до силура).
Так или иначе, но расширение было неизбежно.
Дело в том, что нейроны нуждаются в защите друг от друга, а организм и его иммунная система нуждаются в защите от прожорливости и аффектированной электрохимической активности нейронов.
Соответственно, в образующемся «загущении» разрастается и масса глиальных клеток, которые изолируют нейроны и питают их. Уже одно это обстоятельство троекратно увеличило объёмы нашего «утолщения».
Но это ещё не всё.
Нейроны нуждаются не только в защите друг от друга; в такой же степени они нуждаются и во взаимодействии. Для их контактов тоже требуются значительные анатомические «пространства».
(В спинном мозге их связи легко обеспечиваются протяжённой пучковой структурой самого тяжа.)
В новом «коротеньком» образовании эти связи потребовали объёмного (вертикального и горизонтального) разрастания субстрата, что наметило своеобразную геометрию будущего мозга.
Естественно, вся эта конструкция не могла бы существовать без своей собственной системы кровообращения и ликворотворения. «Мощностей» мягкой и арахноидной оболочки, унаследованных от спинного мозга, естественно, хватить не могло. И этот фактор тоже привёл к существенному размерному разращению тех зачатков мозга, которыми располагали позвоночные на заре палеозоя.
Естественно, эволюция позвоночных не собиралась останавливаться на обострении тактилизации.
Вероятно, со временем её одной стало недостаточно для успешности организма в ту суровую пору; враги были изобретательны, половые партнеры — капризны, а еда — подвижна.
Постепенно наше нейронное «загущение» обзаводится нейропилем — зачатками той самой ретикулярной формации, которая обеспечивает предельную отчётливость связи меж спинным мозгом и нарождающимся головным.
 | |||
 | |||
Илл. 53. Головной отдел древнейшей бесчелюстной рыбы (Heterostraci).
Силур. Прим. 420 млн. лет назад. (По Оленеву)
И вот тут-то уместны и фейерверки и литавры: наконец-то в истории мозга появляется хоть что-то, к чему уже может быть полноценно применена нейроанатомическая латинская номенклатура. В данном случае это будет tractus spinoreticularis и tractus reticulospinalis.
Вот примерно так и образовалась первая фундаментальная часть головного мозга, именуемая ныне metencephalon, задний мозг.
Учитывая ценность данного новообразования и его большой практический смысл для организма, «загущение» постепенно облачалось в защитную костную капсулу.
Происхождение капсулы дискутивно. По одной из версий материалом для неё послужили слившиеся позвонки, а подругой (более разработанной теории) — костно-кожисто-хрящевые пластинки, вроде тех, что защищали головы акантод (первых примитивных рыб).
В этом вопросе «клинки» гипотез (в свое время) скрестились очень впечатляющим образом. Основоположником позвонковой версии был И. В. Гёте (1749-1832), малоизвестной страстью которого была сравнительная анатомия, и который в 1820 написал: «В 1791 году, когда нашёл на песчаном бугре еврейского кладбища в Венеции старый изломанный череп овцы, я заметил, что и личные кости образованы из большого позвонка. Когда при внимательном осмотре мне удалось увидеть постепенность перехода от первой крыловидной кости к решетчатой и её губчатым частям — всё остальное стало для меня ясным». Гете был не одинок, почти одновременно с ним позвонковую версию муссировал иенский профессор Лоренц Окен, а чуть позже и сам сэр Ричард Оуэн, линнеевский кавалер. Противниками этой гипотезы были (естественно, в разное время): Кювье, Райхерт и Т. Г. Хаксли, как раз и породившие кожисто-хрящевую версию. А. Дорн в «Происхождении позвоночных и принципе смены функций» (1875) вообще предлагает рассматривать появление черепа как ответ эволюции на необходимость опоры для язычной и челюстной мускулатуры, что маловероятно, так как согласованная, тонкая и мощная работа челюстей никак не могла появиться раньше зачатка мозга, способного координировать эти сложные движения и вообще сделать их нужными. Более того, последние находки подтвердили, что и многие бесчелюстные уже были снабжены черепами (костными головными щитами: Astraspis, Poraspis, Tolypelepis). Полагаю, настоящий ответ похоронен в кембрийских (или ордовикских) горячих морях и болотах, где жизнь позвоночных обретала свои очертания, способности к сложному поведению и свой физиологический «первоначальный капитал»51.
Вот как описывает эти существа академик Д. Северцов: «Протокраниаты были, по всей вероятности, голыми животными, т.е. их кожа не была покрыта чешуями... а на переднем конце тела, вероятно, из скелетных образований метамерного происхождения, уже образовался зачаточный осевой череп. Этот череп был построен чрезвычайно примитивно, и головной мозг обхватывался им снизу и сбоку; череп состоял из парахордалий, а возможно, также и из трабекул, которые, однако, к черепу ещё не приросли» (Северцов А. Главные направления эволюционного процесса, 1967).

Илл. 54. Сформированный череп костной рыбы
Параллельно с облачением в череп развивалась как новая рецепторика (зрение-обоняние), так и «её» структуры — средний и передний мозг.
В результате всех этих процессов возникла концентрированная, физиологически обеспеченная, анатомически оформленная и защищённая группа нейрональных структур, которая стала интегративной — сперва для более слабых и рассеянных по организму частей нервной системы, а затем и для всех позднеэволюционных образований головного мозга.
CAPUT XII
Личность организма. Стереотип. Моральная мутация.
Возраст личности. Терапсиды. Всевластие среды.
Прогностизм. Труд, мастурбация и ложь.
Эксперимент Дж. Олдса. Межножковое ядро.
(Дудочка крысолова». Намёк белой крысы. Боль.
Атавизмы разума. Иприт. Рука. Безразличие нейронов.
Этапность цереброгенеза и локализация функций.
Полагаю, понятие «личность» сложно воспринимается в том качестве, в каком я употребляю его в данном исследовании.
Дело в том, что религия, философия и литература наделяют «личностью» исключительно представителей вида homo и декларируют, что именно её наличие обособляет человека от всего остального животного мира.
Этому есть простое объяснение: религия, философия и литература формировались со значительным опережением естественнонаучных знаний, находясь под сильным влиянием абсолютного антропоцентризма, в т.ч. представлений о «высшести» человека, «центральности» его роли в мироздании и так далее.
Напомню, что в тот период, когда создавалась религиознофилософская легенда о «личности», не было знаний о реальной продолжительности истории homo. О том, что (примерно) 100 (на тот момент) поколениям людей, зафиксированной истории — предшествовали (примерно) 80000 поколений людей, бывших обычными животными.
Не существовало никаких хотя бы приблизительных представлений о функциях мозга и нервной системы, о сравнительной нейрофизиологии и эволюции. Разумеется, не было даже догадок о том, что «личность», или «биологическая индивидуальность», есть обязательная функция мозга на любом уровне его развития, которая и позволяет существу самоосознавать себя и выстраивать отношения со своим организмом как с безусловной собственностью.
Но рожденная в ту мифотворческую пору трактовка «личности» дожила в неприкосновенности и до наших дней.
К XIX веку религия, литература и философия окончательно «раскрасили» её в пафосно-драматические цвета и, сакрализировав, передали XXI столетию как уникальную ценность и стержень всей земной истории и культуры.
Любопытно, что и сейчас «личность» предполагается не у вида homo во всей его естественноисторической полноте, а только у тех особей, что появились на свет, когда человек из дикого животного преобразился в существо, живущее по правилам социализации. (А этот период, как мы помним, составляет лишь 0,23-0,25% от общей продолжительности истории того homo, что перешёл «рубикон Валлуа»).
У данного факта есть своя причина: мифологизированное понятие «личность», со всей её атрибутикой, вроде «свободы воли»52, «греха», «стыда», «совести», «подвига» и так далее, никак не увязывается с образом обычного стайного животного, промышлявшего поиском падали и съедобных кореньев, хотя, как мы знаем, головной мозг у примерно 80000 «диких поколений» homo не имел существенных отличий от мозга последующих им 200 «социальных» поколений.
Исходя из вышесказанного человек и не мог унаследовать иную «личность», кроме той, что формировалась у всей его предковой цепочки за 500000000 лет и почти 2000000 лет демонстрировалась ранними homo.
Социальные игры и ритуалы, язык и интеллект могли лишь декорировать «личность» в соответствии с модами и тенденциями столетий, упорно научая животное homo играть «человека»; скорее играть тот образ, который из ограничений, грёз и традиций «вылепили» мифология и законы.
Прекрасно понимаю, до какой степени некорректно противопоставлять друг другу мифологию и биологическую реальность. Победитель в этой схватке предопределён, но у меня не было ни малейшего желания устраивать избиение младенцев; я же не виноват, что подходящий мне ёмкий термин (личность) столь плотно оброс культурологическими ассоциациями и фантазиями.
Классическая эволюционная нейрофизиология на данную тему высказалась аккуратно, но твёрдо ещё в середине XX века: «Те формы поведения, которыми отличались наши предки, жившие миллионы лет назад, все в нас гнездятся и при определенных условиях проявляются» (Орбели Л. А. Об эволюционном принципе в физиологии, 1961).
Следует отметить и факт принципиальной невозможности провести отчётливую демаркационную линию меж мозгом «диким» и «социализированным»: «Главная трудность этой проблемы состоит в том, чтобы понять, в какой степени человек отрывается от животного мира. Проблема осложняется необходимостью решить вопрос, который всегда является камнем преткновения — о соотношении биологического и социального факторов в развитии человека... Нам представляется, что этот вопрос также нужно решать с эволюционной точки зрения» (Карамян А. И., акад. И. М. Сеченов и эволюционная нейрофизиология, 1980).
Любая же попытка решения этого вопроса с «эволюционной точки зрения» неминуемо вынудит нас вернуться к простому пониманию того, что «все современные формы жизни являются прямыми потомками тех, которые жили задолго до кембрийской эпохи» (Darwin) и вынудит искать закладные, исходные черты «личности» во времена зарождения органической жизни.
Но говорить о специфике «докембрийской» личности в высшей степени сложно, так как наши представления о нервной системе организмов той эпохи остаются строго теоретическими. (У нас немало красноречивых компонентов, дающих право на обоснованные догадки и аккуратные выводы, но ещё нет и, вероятно, уже не будет связной и целой картины, т.к. нет аутентичного «препарата». Стиль и состояние всех палеозоологических находок как эдиакарской, так и предшествующих ей эпох не дают никакого основания предполагать, что такой «препарат» появится.)
Понятно, что магистральные черты «личности» неизменны и обусловлены (в первую очередь) «конфликтными» свойствами нейрона, что первый поворотный пункт в её истории — это появление протоконодонтов и, соответственно, «первая кровь», но я бы предпочёл руководствоваться более полной, однозначной, а главное — проверяемой сегодня фактурой, пусть даже и отнесённой от вендской эпохи на несколько сотен миллионов лет.
Такая существует.
Мы можем взять за очень (очень!) условный второй поворотный пункт истории «личности» появление терапсид (Therapsida) или пеликозавров (Pelycosauria); рептилий, не только обречённых на достаточно сложное поведение, но и уже способных на него53 (илл. 55).
И пеликозавры (букв. парусные ящеры), и терапсиды относятся к подклассу Synapsida класса Reptilia, их явление свершилось спустя почти 300 миллионов лет после эдиакарской эпохи, в т.н. пермский период (298-251 млн. лет назад). Палеозоологи до сих пор дискутируют о границе меж этими двумя разновидностями отряда синапсид (Synapsida), но предлагаю им и оставить эти диспуты.
Нам вполне достаточно того, что это были зверозубые пресмыкающиеся, уже имевшие зрение, обоняние и чудесный череп, прямо свидетельствующий о наличии у них полностью оформленного ствола головного мозга и ещё нескольких важнейших структур, включая, естественно, лимбическую систему, т.е. амигдалу, гиппокамп, сосцевидное тело, гипоталамус и так далее.

Илл. 55. Пеликозавр
По черепу лемурозавра, известному как NMQR-1702 и очень характерному для терапсид (на илл. 56), легко «прочитывается», что из оральной части спинного мозга (вполне штатным образом, т.е. под влиянием VIII и X нервов) уже окончательно сложился задний мозг (metencephalon).
Очевидная зрячесть звероящера — гарантирует наличие среднего мозга, а развитая (до полной анатомической оформленности ноздрей) обонятельная рецепторика — предполагает неизбежность наличия развитого переднего мозга.
Понятно, что меж всеми этими отделами мозга установлена функциональная и структурная связь, так как цефализация всегда идёт по одной и той же схеме: «При формировании новой части мозга к ней протягиваются рецепторные пути от всех низлежащих формаций, а возникающие в ней эффекторные пути тянутся к эффекторным путям низлежащих формаций» (Сепп Е. История развития нервной системы позвоночных, 1959).
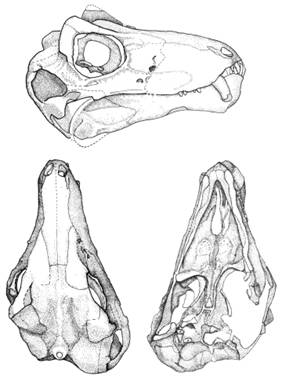
Илл. 56. Череп Lemurosaurus pricei
Явное фактическое наличие этих трёх структур, их несомненная развитость и связанность меж собой позволяют сделать вывод о том, что головной мозг Therapsida располагал как сложившейся ретикулярной формацией, выполняющей интегративную роль, так и лимбической системой (или её крайне существенным зачатком), генерирующей (в том числе) весь необходимый набор агрессий.
Данную точку зрения можно с определенной уверенностью опереть как на параметры черепа терапсиды, так и на ту полноту знаний о строении и специфике мозга современных рептилий, которой мы располагаем на данное время:
«Передний мозг рептилий — это конечное звено многих анализаторных систем» (Сергеев Б. Эволюция ассоциативных временных связей, 1967).
(Раскрыть эту цитату можно следующим образом: у рептилий, сравнительно с амфибиями, усложнилось строение как паллиума, так и неопаллиума и стратиума. Обогатились и существенно «разнеслись» все афферентные связи.)
«Изучение мозга рептилий показывает, что основные зоны коры выражены у них столь же чётко, а частью (переходные зоны) и более чётко, чем у млекопитающих, что кора больших полушарий при всей огромной дислокации этих зон в процессе филогенеза, обусловленной ростом новой коры, построена в обоих классах по единому плану» (Филимонов И. Сравнительная анатомия большого мозга рептилий, 1963).
Более того, «у рептилий впервые в ряду позвоночных оказывается возможным образование временных связей типа ассоциации» (Карамян А. Функциональная эволюция мозга позвоночных, 1970).
Полагаю, что данные постулаты А. Карамяна, И. Филимонова, Б. Сергеева безупречны, но нуждаются в небольшой нейроанатомической расшифровке.
Итак.
Вкратце рассмотрим мозг ящерицы Lacerta agilis, варана Varanus griseus и аллигатора Alligator mississippiensis.
Что мы видим?
Во всех трёх случаях наличествуют приметы очевидного «конструкционного» сходства: чётко выделен септальный бугор (colliculus septalis); полушария расширены в заднем отделе, а кпереди суживаются и переходят в обонятельный тракт; присутствуют (в разной степени отчётливости) циркулярные, септостриатные, ринальные, эндоринальные и латеральные борозды, желудочки, обонятельные луковицы и тракты, боковые ямки (fovea lateralis), ядра дорсального стриатума, амигдалярные комплексы, палеокортекс, археокортекс, лимбика и так далее. (Наблюдения проведены мною по методу И. Филимонова (1963) и сверены с Капперсом (1921), Герриком (1956), Адиссоном (1915), Ризе (1928), Лангуорти (1937), Ретциусом (1898).)
Исходя из вышесказанного мозг рептилий многофункционален, сложен и содержит в себе все структурные компоненты, которые обеспечивают сознание, ассоциации и адресации.
Это мозг полноценного и крайне эмоционального существа, имеющего весь набор зрелых агрессий и готовность их реализовывать. То, что мы называем «крайней жестокостью», всего лишь свидетельство адаптативности, способности к адекватной оценке среды и, в известной степени, — недостатков памяти и сознания, обусловленных некоторым несовершенством гиппокампальных структур и рецепторов.
Есть все основания предполагать, что очерченная выше схема полностью применима и к терапсидам пермского периода.
Разумеется, возможны сомнения в том, что за 250 миллионов лет церебральные структуры рептилий остались неизменными, а результаты исследований XIX и XX веков применимы к первым звероящерам. Эти сомнения, конечно, оправданы, но вероятность того, что существенных изменений мозг рептилий не претерпел, всё же неизмеримо выше любых предположений о его качественных метаморфозах.
Дело в том, что современное состояние мозга рептилий являет собой одновременно и некий эволюционный «апогей» (для подобного существа), и то состояние, «ниже которого» никакое адекватное поведение (для организма подобного типа) вообще невозможно.
Иными словами, столь сложный организм и не может управляться менее развитым мозгом.
Впрочем, ничего экстраординарного в факте неизменности головного мозга в течение столь длительного времени нет.
Прекрасной иллюстрацией данного положения может служить Astacus fluviatilis (речной рак).
Рассмотрев рака, мы увидим, что его общая морфология не допускает «вычитания» ни единого из компонентов его мозга (надглоточного, комиссурального, подглоточного ганглиев и окологлоточной коннективы).
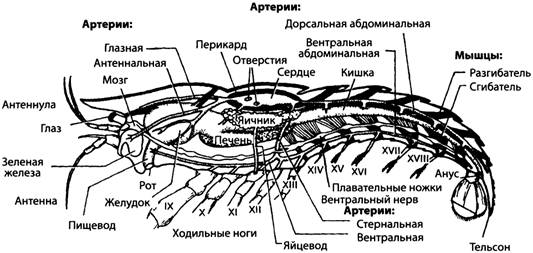
Илл. 57. Главные системы органов речного рака
Любая воображаемая (или реальная) попытка упрощения этого мозга сделает организм такого типа неуправляемым: поведение будет неточным, физиология — неконтролируемой, движение — невозможным. Неизменность же общей морфологии Astacus с кембрийского периода подтверждена многочисленными палеозоологическими находками и исследованиями Т. Г. Хаксли «Рак. Введение в изучение зоологии» (1900), Я. Бирштейна «Десятиногие» (Основы палеонтологии, 1960), «The Physiology of Crustacea» (1961, Vol. 2), Ж. Шуранова, Ю. Бурмистрова «Нейрофизиология речного рака» (1988).
Возможно, здесь не лишним будет напоминание о тех особенностях развития и существования жизни, которые прекрасно были сформулированы академиком А. Северцовым: «Факт благополучного выживания (и даже известного биологического процветания) крайне примитивных форм, остатков фауны давно прошедших времён является крайне странным. Мы уже указали на некоторые примеры такого выживания (круглоротые, эласмобранхии), но едва ли многие читатели представляют себе, насколько это явление оказывается закономерным и распространённым в природе... Современная фауна состоит, таким образом, из форм животных, принадлежащих по высоте и характеру своей организации к самым различным по времени эпохам существования земного шара» (Северцов А. Главные направления эволюционного процесса, 1967).
Впрочем, вернёмся к черепу терапсиды.
Повторяю, череп NMQR-1702 достаточно типичен, прекрасно изучен (Sidor C.A., Welman J. A Second Specimen of Lemurosaurus pricei. Therapsida: Burnetiamorpha, 2003), в том числе и в вопросе принципиальной сопоставимости с черепами других синапсид пермского и триасового периодов (горгонопсиды, буллоцефалуса, лобалопекса, диметродона, доцинодона и так далее), чтобы служить эталоном и давать право на некоторые обобщения.
Обобщим.
Головной мозг звероящеров уже достаточно совершенен.
Основные структуры, обеспечивающие сознание, эмоции, самоидентификацию (личность и её инструментарий), сложное поведение — уже сформировались.
Остались лишь мелкие дорисовки, которые завершатся через 200 миллионов лет, когда млекопитающие потомки терапсид сменят динозавров на сцене эволюционного театра.
Разумеется, личностные особенности первых звероящеров были лишь одним из этапов развития этой генеральной функции мозга, но отнюдь не её «закладным камнем», не фундаментом и не первоосновой. Самые начальные характеристики как были, так и остаются во тьме архея и протерозоя.
Тем не менее биологическая личность homo является прямым продолжением личностных особенностей в том числе и зверозубых существ пермского периода. Это особенно хорошо видно на примере как гомологичности ядер ретикулярной формации, лимбической системы (см. гл. II), так и при сопоставлении иных структур мозга: «У высших млекопитающих, особенно у человека, зрительный бугор развит очень сильно в связи со значительным развитием мозговой коры. Его функциональная и структурная дифференцировка очень детальна. Однако основная схема строения и связей остаётся такой, какой она начала формироваться на уровне амфибий и сложилась у рептилий» (Сепп Е. История развития нервной системы позвоночных, 1959).
Млекопитание, плацентарность, обогащение рецепторики внесли в эти особенности свои очень существенные коррективы, но никак не принципиальные изменения.
В контексте нашего исследования стоит особо отметить развитие V и VII черепных нервов (млекопитающие унаследовали их опять-таки от рептилий, правда, в весьма скромном виде).
Именно n. trigeminus и n. facialis явились организаторами мимического языка млекопитающих, значительно более универсального, чем запаховый, позный, пластический, экскреторный и прочие языки. Трудно сказать, насколько именно он универсален в масштабе всего класса (Mammalia), но в рамках отрядов и семейств, а тем более родов и видов его универсальность несомненна.