Сегодняшняя большая настойчивость в «потягивании» «распускает» и всё «плетение» как ранних, так и поздних доктрин о конструкции мозга.
(Ознакомившись с «пророческой» ремаркой анатома Чарльза Белла, уже совсем сложно избавиться от подозрения в том, что суть всей эволюции — это, прежде всего, история мозга и его трансформаций. А всё многоразличие черепов, глазниц, карапаксов, костно-хрящевых щитков — это лишь маскарадные наряды мозга, сменяемые им в строгом соответствии с «модами» и потребностями геологических или иных эпох.)
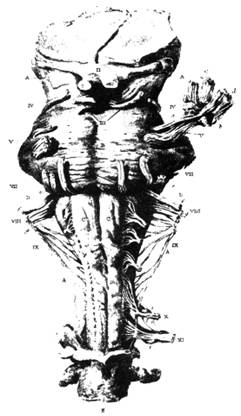
Илл. 41. Таблица из труда Ч. Белла о нервной системе, демонстрирующая ствол мозга
Впрочем, вернёмся к нашей теме.
Надо отметить научную осторожность, которую демонстрируют все без исключения исследователи, когда речь заходит о роли ретикулярной формации в процессах образования мышления или разума.
Их легко понять, тут действительно возникает существенная сложность, требующая «дерзости и обобщений». Понятно, что любой из крупных учёных, кто попытался бы сформулировать положения ретикулярной формации в этом аспекте, рискнул бы научным именем.
Мы все помним, какой шок вызвало невинное заявление д-ра М. Шайбель на Детройтском конгрессе 1957 года.
Тогда сам сэр Дж. Джефферсон вынужден был внести коррективы в свой доклад «Ретикулярная формация и клиническая неврология», чтобы охарактеризовать реплику д-ра Шайбеля как «невероятную»: «Д-р Шайбель, после того как он представил свои прекрасные гистологические препараты, выступил с просто невероятным заявлением, что “это” (ретикулярная формация) и есть центральная нервная система».
Полагаю, что есть необходимость расшифровывать смысл характеристики «невероятная» из уст сэра Джеффри Джефферсона (1913-1961), радикального академиста, клинициста и крупнейшего авторитета нейрологии.
Сэр Джеффри был знаменит безупречным лексиконом и сверхделикатностью. Данная реплика была для него крайним лексическим пределом при выражении недоумения и гнева.
(Полагаю, все вынесенные за XX век смертные приговоры не могли бы конкурировать в убийственности с этой характеристикой.)
Я в лучшем положении, чем д-р М. Шайбель. У меня нет никакой научной репутации, и рисковать мне нечем. (Ita et sir Jeffri pridem mortuus est. — Да и сэр Джеффри давно мёртв.)
Полагаю, что если попытаться кратко сформулировать суть теории ретикулярной формации применительно к таким явлениям, как мышление и разум, то мы увидим следующее: ствол мозга homo, включающий продолговатый мозг, мост и средний мозг, — содержит в себе древнюю структуру, управляющую мозгом и использующую все поздние его формации как свои инструменты. Необходимость в столь сложном инструментарии вызвана исключительной сложностью организма, которому надо обеспечить выживание.
Это управление, вероятно, является не прямым, а строго опосредованным, через активацию, возбуждение и торможение различных, в том числе и корковых зон.
(Древние стволовые структуры используют кору точно так же, как они используют зрительную систему или другие анализаторы.)
По ретикуло-кортикальным путям идёт активация (или торможение) участков коры (проекционных или ассоциативных центров), там происходит акт анализа, а кортико-ретикулярные связи обеспечивают передачу в ствол мозга его сущностной части.
Судя по плотности нейронов в тех ядрах, что вовлечены в прямую и обратную связь с корой (латеральное ретикулярное ядро, парамедианное ретикулярное ядро, ретикулярное ядро покрышки моста), «сеточке» предлагается строгая, точная выжимка, некий информационный концентрат. (Плотность нейронов колеблется в различных ядрах, но она везде невелика. Достаточно сказать, что из всей массы нейронов ретикулярной формации, непосредственно в ядрах — заключено едва 5-7 %.)
Косвенным, но сильным подтверждением интегративной роли ретикулярной формации служит и тот, уже ставший безусловным, факт, что именно стволовые структуры являются (образно выражаясь) «повелителем сна и пробуждения».
Ненадолго остановимся на понятии «сон».
Не будем забывать о литературно-бытовом характере этого термина, для нас, опять-таки, ничего не означающем. Такого понятия, как «сон», строго говоря, не существует. Оставим его романистам. Мы можем говорить лишь об активности части коры, от минимальной (т.н. сна) до максимальной.
(Естественно, т.н. сон имеет несколько фаз и вариаций, но я здесь рассматриваю не частности, а принцип.)
Неслучайно, первое, с чем «встречается» любое исследование ретикулярной формации, это её возможность как провоцировать сон, так и прекращать его. В переводе на нейрофизиологические понятия это не означает ровным счётом ничего больше, как способность регулировать активность коры головного мозга. От максимальной до минимальной.
И ретикуло-кортикальные и кортико-ретикулярные связи обстоятельно исследованы и доказаны трудами Дежерина (1901), Занда (1903), Симпсона и Джолии (1907), Метлера (иссл. 1935-1947), Левина (1936), Верхарта и Кеннарда (1940), Минклера и Клемма (1944), Комса (1949), Эсколара (1950), Крига (1954), Джаспера, Аймоне-Марсана, Столла (1952), Бремера и Терзуоло (1954), Френча, Эрнандес-Пеона и Ливингстона (1955) et cetera.
Разумеется, есть много пока непонятного и требующего кропотливых локальных исследований.
К примеру, Дж. Ф. Росси и А. Бродал пришли к выводу, что «волокна из самых различных частей коры головного мозга оканчиваются в одних и тех же областях ретикулярной формации» (Rossi G. F., 1957; Brodal A., 1958).
Причина этого явления неясна.
Г. Кейперсу удалось установить, «что корково-ретикулярные пути организованы примерно одинаково у всех животных, включая человека» (Kuypers Н. G.J., 1958), что является, полагаю, рискованным заявлением, так как мы явно не набираем ту статистическую массу (основанную на грамотных экспериментах и анализах), которая позволяла бы делать столь широкие обобщения.
Повторяю, теория не претендует на истину, вполне возможно, что она так же ошибочна, как и многие другие, но её важнейшая (по моему мнению) часть, т.н. центрэнцефалическая, тем не менее, останется в силе, так как заподозрить головной мозг в равноправии всех его структур категорически невозможно.
К слову, есть и ещё одна важная причина, по которой я из множества теорий нейрофизиологии выбрал, как основной инструмент, именно «ретикулярную», но об этом чуть позже.
Конечно, принцип некой иерархии неоспорим. Он отмечен (явно или туманно) в любой из гипотез, начиная с декартовской, где загадочное «главенство» над мозгом отдано эпифизу.
Везалиус и Велизиус полагали, что управляющим органом мозга является полосатое тело corpus striatum (т.е. хвостатое ядро, скорлупа, бледный шар). Этот выбор никак особо не мотивировался (равно как и выбор Декарта), поскольку имелся в виду (строго говоря) не интегративный, или командующий, центр мозга, а лишь место обитания «души», у которой были все управленческие полномочия.
Чуть позже все руководящие функции были приписаны коре. Считалось, что коль скоро именно кора генерирует «наивысшие» особенности человека, то ей и принадлежит право управления его мозгом.
Полагаю, такой вывод поспешен, а причина его лишь в слепой сакрализации интеллекта, в нежелании определиться с его местом и эволюционным значением.
(Стоит отметить, что ранее объектом такой слепой сакрализации была «душа», и ей тоже приписывались генеральные функции.)
Безусловно, кора млекопитающих имеет уникальные свойства, но здесь будет уместна следующая аналогия:
Прибор ночного видения, сверхдорогой и сверхточный, в известном смысле слова руководит действиями снайпера, желающего произвести прицельный выстрел в полной темноте. В этой ситуации зависимость снайпера от прибора огромна, влияние прибора на принятие решений — безусловно. И тем не менее этот прекрасный прибор остаётся лишь инструментом снайпера, а не становится управляющей силой в понятном смысле этого слова.
Аналогия мне представляется грубоватой, но корректной, так как интегративные структуры мозга действительно находятся в «полной темноте» сформировавшего их палеозоя, и им требуются совершенные анализаторы для принятия верных решений (илл. 42).
(Полагаю, что в суть теории ретикулярной формации можно заложить примерно такое понимание соотношения коры и древних структур.)
Конечно, это по существу прямая декларация зависимости коры со всем её проекционно-ассоциативным аппаратом, т.е. с разумом и мышлением, от первородных «примитивных» образований мозга.
(Но следует помнить, что не всякая зависимость лишает определённой свободы.)
К слову, ещё одним и, как я полагаю, крайне весомым аргументом «подчинённости», «инструментальности» коры — будет и упоминание о том факте, что такой важнейший фактор формирования множества жизнеорганизующих процессов, как боль — коре неведом. «Боль не имеет своего представительства в новой коре больших полушарий» (Бериташвили И. С. Структуры и функции коры большого мозга, 1969).
Итак, используя все эти выкладки, попробуем напрямую обозначить основной вопрос: что в головном мозге homo является управляющим и определяющим — природное «животное» или искусственное «интеллектуальное»?
Рассмотрим этот вопрос ещё тщательнее.
Да, если мы принимаем «ретикулярную теорию» как основу представлений о механизмах мозга, то тем самым мы признаём полную и прямую зависимость аналитических потенциалов коры от «примитивных и слепых» древних формаций мозга, по сути, единых для всех видов животных, структурировавшихся во времена палеозоя примерно триста миллионов лет назад.
Причём следует признать, что эти формации активируют проекционные и ассоциативные участки неокортекса вне всякой связи с тем, какой конечный продукт будет результатом этой активации.
(Вернее, можно было бы сказать, что вполне возможен и мало предсказуем некий добавочный эффект этой активации, помимо основного, биологического эффекта.)
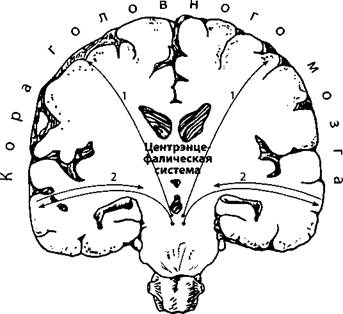
Илл. 42. Схема связи коры головного мозга с центрэнцефалической системой
Поясняю.
Для коры «мауглеоида», homo erectus или Эйнштейна сработают одни и те же механизмы активации-торможения, идущие от стволовых структур.
Для примера — самое простое действие.
Предложим всем трём нашим персонажам взглянуть на ночное небо. (Эйнштейну, «мауглеоиду» и homo erectus.)
Полагаю, всем троим будет достаточно обычного жеста указания на него, никакой вербальной конкретизации не потребуется. Объект настолько прост и велик, что промахнуться взглядом невозможно.
Далее всё будет происходить почти идентично.
У всех троих персонажей последует согласованная краткая работа нескольких мышц шеи, затем — пяти мимических мышц, синхронно с ними сработают четыре из шести мышц глазного яблока, приподняв его вверх, и сетчатка глаза примет пропущенную через хрусталик «картинку».
Перевернутая «картинка ночного неба» по n. opticus отправится в латеральное коленчатое тело, оттуда — к шпорной борозде, в проекционный центр зрения. Примечательно и крайне важно, что в проекционном центре перевернутость картинки не исчезнет.
«Из-за перевернутой картины, образуемой на сетчатке при помощи хрусталика, верхнее зрительное поле проецируется на нижнюю область сетчатки и передаётся в область V1, расположенную ниже шпорной борозды; нижнее зрительное поле проецируется над шпорной бороздой» (Nicholls J. G., Martin A. R., Wallace В. G., Fuchs Р. A. From Neuron to Brain, 2006).
Далее картинка проследует в ассоциативный центр, на дорсальную поверхность затылочной доли, где, собственно, и «перевернется» (илл. 43).
(Чрезвычайная важность этого простого обстоятельства заключается в том, что факт переворота картинки лишь на самом финальном этапе, в высшем корковом ассоциативном центре, есть наилучшее свидетельство глубочайшей древности этого центра и того, что архитектура мозга была завершена значительно ранее, чем это предполагается.)
Сам переворот — несложное действие, но он происходит только здесь. Лишь попав в «ассоциативный центр», зрительная информация становится полноценной, т.е. позволяющей принимать верные поведенческие решения. И это касается как homo, так и любого другого животного любой степени древности. Следовательно, этот центр — обязательное условие полноценности любого мозга, а не только его «позднеэволюционной модификации», и существует он не в силу того, что поздний homo начал грузить его сложными интеллектуальными ассоциациями и образами.
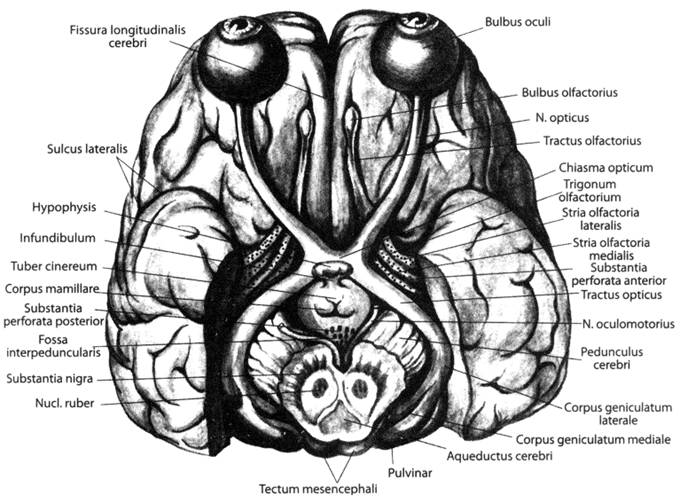
Илл. 43. Зрительные нервы и тракты (по Синельникову)
(К слову, как и в случае с зонами Брока и Вернике, мы в очередной раз убеждаемся в изначальной эволюционной заготовленности зон коры под сложные функции.)
Впрочем, мы чуть отвлеклись от нашей троицы. От Эйнштейна, homo erectus и «мауглеоида», ну скажем, 1925 года рождения. (Или любого другого, но находящегося в рамках условной «современности».)
Для мозга всех троих процесс восприятия картинки ночного неба, её прохождения по церебральным структурам и прибытия в ассоциативный центр — абсолютно идентичен. (Другого просто не предусмотрено физиологией мозга.)
Вполне вероятно, что и ретикулярная формация будет одинаково активировать структуры, по которым «проходит» картинка (от ганглиозных клеток сетчатки до дорсальной поверхности затылочной доли). Также вероятно и, более того, неизбежно, что для оценки увиденного и правильной реакции на него будут активированы и все остальные корковые зоны.
При примерно равном количестве нейронов в ассоциативных центрах, их анатомическом подобии, идентичности функций и одинаковом возбудителе — у всех этих трёх представителей homo и продукт активации коры должен был бы (по крайней мере) сходствовать. Однако это категорически не так.
При взгляде на ночное небо продуктом кортексной активности мозга Эйнштейна будут нюансы представлений об искривлении пространства или правки в модель космологической динамики. (Si, vero, poterit se demovere a cogitationibus de gluteo maximo Elzae Levental. — Если, конечно, сможет отвлечься от размышлений о большой ягодичной [мышце] Эльзы Левенталь)
Продуктом кортексной активности мозга homo erectus и «мауглеоида», несмотря на разделяющий их миллион лет, будет лишь ощущение наличия над их головами темноты с яркими точками. (И это в самом лучшем случае, полагаю, что даже подобное предположение фантастично).
Скорее всего, сам «факт неба» и его звёздности не вызовет ни интереса, ни самой минимальной реакции. Чёрное небо не опасно само по себе, оно не съедобно и не означает ровным счётом ничего, кроме утраты предметами и существами той отчётливости, которая возможна при ярком свете.
Но ретикулярная формация ответ на свой вопрос, ради получения которого она активировала различные области мозга, всё же получит; из двух реальностей, тёмной и светлой — в наличии тёмная, более опасная. Естественно, этот «ответ» будет гораздо нюансированнее, сложнее и точнее, чем тот, что можно вместить в слово «опасная», но примерный смысл будет, вероятно, именно таким.
Простой пример: даже ощущение опасности — и то двояко. Вместе с концентрацией внешней угрозы, генерируемой «тёмным миром вокруг», у homo проявляется и физиологическое знание повышения его собственной угрозы для этого «мира», так как мрак и ему даёт некоторые повышенные шансы убить, совокупиться, украсть и так далее.
На «символ темноты» отзовётся механизм агрессий. Он спровоцирует и у «мауглеоида», и у homo erectus страх или настороженность, отрефлексирует их на резкую оглядку по сторонам, мобилизует обоняние, слух и через ретикуло-спинальный тракт активирует всю мышечную систему.
Строго биологическая (важнейшая) задача будет выполнена.
(Стоит отметить, что данная схема (если она верна) будет справедлива применительно как к homo erectus, так и к мауглеоиду, несмотря на разделяющий их миллион лет.)
Но вот глубокая прозрачная чернота неба и светящиеся в ней точки звезд — не будут и не могут иметь для них никакого смысла.
По одной простой причине. Отсутствуют номинации, ничто не имеет имени. Та часть реальности, что не опасна, не съедобна, не вожделенна, т.е. находится за пределами «круга понятного», круга, очерченного инстинктами, остаётся безымянной (как в целом, так и в мельчайших частностях), её бесконечное многообразие бессмысленно... и неинтересно.
Сознание мауглеоида и homo erectus строго сиюминутно, причём у этой сиюминутности нет ни начала, ни продолжения. Зрительные и иные ассоциации, разумеется, возможны, но они имеют стихийнохаотичную структуру и плохо управляемы, так как отсутствует их классификатор в виде тех «меток», или номинаций, которые могли бы позволить ими пользоваться (управлять).
Как видим, разница продуктов возбуждения коры, конечно, не огромна, но любопытна.
Даже не признавая за интеллектом никакого особого эволюционного значения, даже понимая, что это лишь некая «погремушка мозга», побочный эффект возбуждения коры, тем не менее, мы вынуждены констатировать наличие неявного фактора, который и определяет его наличие или отсутствие.
(К слову, я выбрал для примера Эйнштейна исключительно по сопоставимости массы его мозга с типовой массой мозга палеоантропа (1222 г) и средними значениями обследованных «мауглеоидов». Масса не имеет почти никакого значения, но в данном случае это просто выполнение правил подобных экспериментов.)
Какие физиологические процессы могут быть в основе этой разницы? Если исходить из догм, то разница «продукта» должна быть обусловлена разницей стуктур или процессов. Понятно, что в нашем случае говорить о «разнице» невозможно, следовательно, остаются «различия».
Поищем их физиологическое обоснование. Круг поисков невелик.
Первый претендент на обеспечение этого различия — интенсивность иррадиации возбуждения-торможения.
У иррадиации есть прекрасное, чёткое определение, данное ей И. П. Павловым: «Раздражение, пришедшее в большие полушария, сначала разливается, иррадиирует, а затем собирается, концентрируется». Интенсивность иррадиации может зависеть от силы активации (возбуждения) и от особых свойств клеток.
Но по условиям нашего эксперимента, сила возбуждения (по крайней мере формальная) одинакова.
Тогда посмотрим клетки.
Опять-таки, обратившись к догматике, мы можем выяснить, что в определённых клетках нервной системы может быть несколько ядер, и что размеры ядрышек в этих ядрах могут увеличиваться и уменьшаться под воздействием внешних факторов. Об этом свидетельствуют апологеты радикального академизма: Ж. Эдштром, С. Блинков, И. Гайворонский, И. Глезер, О. Фогт et cetera.
Но тот же И. Гайворонский говорит лишь о клетках вегетативной НС, что особо и подчеркивает: «В нервных клетках вегетативной нервной системы может встречаться по 2-3 ядра. Количество ядрышек в ядре также составляет от одного до трёх. Увеличение числа ядрышек и их объёма свидетельствует об усилении функциональной активной нейрона» (Проф. И. Гайворонский, 2007).
Говоря о клеточных метаморфозах, С. Блинков и И. Глезер подчёркивают, что речь идёт лишь о строго моторных клетках. «Ядрышко в моторных клетках заметно увеличивается в зависимости от функциональной двигательной нагрузки. Увеличение размера ядрышка нервных клеток продолжается до второй половины жизни человека (30 лет)» (Блинков С., Глезер И., 1964 по данным Edstrom 1,1957).
По данным О. Фогт, Ц. Фогт, «с началом старения организма ядрышко постепенно уменьшается и даже может совершенно исчезнуть» (Vogt О., Vogt С, иссл. 1945-1947).
В противовес этим академическим догмам есть столь же академическое мнение творца нейронной теории Сантьяго Рамон-и-Кахаля (1852-1934), которое отчасти конфликтует с вышеприведёнными утверждениями: «На основании многочисленных сравнительных исследований утверждается, что размер тела нервной клетки и диаметр аксона не связаны с физиологической специализацией, а пропорциональны богатству и распространению концевых разветвлений, и следовательно, обилию и многообразию связей» (Кахаль С. Р. Общие рассуждения о морфологии нервной клетки, 1894).
Впрочем, как и полагается гению, в 1904 году Кахаль с легкостью опровергает себя самого, догматизировав в основном труде своей жизни следующее: «Это и многие другие факты учат нас, что морфология нервных клеток управляется не имманентными (внутренне присущими) и фатальными (неизбежными) факторами, передающимися по наследству, как считали некоторые авторы, а зависит полностью от текущих физических и химических условий окружающей среды» (Cajal Santiago Ramon у. Textura del Sistema Nervioso del Hombre у de Los Vertebrados, 1904).
За последнее время ни ревизии, ни примирения всех этих в высшей степени противоречивых догм не произошло.
Наступило время лаконичных обтекаемых формулировок, авторы которых уклоняются от оценки возможности увеличения-уменьшения нейроцитов, их деления, их зависимости от любых факторов, ограничиваясь крайне корректным и почти ничего не означающим заявлением: «Нейроны отличаются от большинства клеток ещё и тем, что за небольшим исключением, не могут делиться» (Nicholls J. G., Martin A. R., Wallace В. G., Fuchs Р. A. From Neuron to Brain, 2006).
(Под «небольшим исключением», вероятно, имеются в виду постоянно обновляющиеся нейроны гиппокампа и обонятельной луковицы (J. Altman, 1965; D. R. Kornack, 1999)).
Таким образом, мы видим, что практически все школы оставляют за размерами и объёмом нейронов право на некоторую неопределённую вариативность. Минимальную или вовсе ничтожную.
Это и нам даёт право усреднить объёмы нейронов, допустив, что коль скоро возможна вариативность в сторону увеличения под влиянием «физических или химических условий», также она возможна и в сторону уменьшения.
Итогом всё равно будет признание малой существенности фактора вариативности нейронов в «иррадиационном вопросе».
Возможно, я не учёл всего, и некоторое влияние фактора объёма отдельных нейронов всё же существует. Но оно гарантированно не может быть глобальным.
Уменьшение-увеличение интенсивности иррадиации могло бы повлечь задержку оценок, ослабление ассоциаций, затруднения номинаций, упростило бы выводы и аналогии.
Но если бы дело было лишь в этом факторе, то homo erectus и «мауглеоид» сгенерировали бы более простые понимания природы звёздного неба, чем Эйнштейн, но это всё равно были бы некие «понимания».
Мы можем продолжить поиск нейрофизиологических различий, которыми можно было бы объяснить разницу конечного «продукта» работы мозга наших трёх героев, но, полагаю, что любой фактор, к которому мы обратимся, от генного до плотности синаптических связей меж нейронами, не окажется настолько глобальным, чтобы объяснить такую разницу в восприятии ночного неба.
Разумеется, подбор персонажей для нашего теоретического эксперимента продуман и неслучаен.
И homo erectus, и мауглеоид — это прекрасные образчики, по сути, самых обычных животных, но животных, имеющих полную церебральную и иную анатомическую сопоставимость или даже идентичность с поздним homo, к примеру, с Эйнштейном.
(Если по homo erectus ещё возможны дискуссии о полной идентичности его мозга и иных параметров мозгу и иным параметрам современного человека (позднего homo), то с мауглеоидом ситуация, вероятно, бесспорна.)
Естественно, остаётся и ещё одна, самая последняя инстанция, куда можно было бы обратиться за разгадкой получившегося у нас парадокса; к нейрохимическим схемам происходящего в клетках коры.
Возможно, здесь, под двуконтурной мембраной, в цитоплазме нейрона, у Эйнштейна свершаются процессы иные, чем у мауглеоида?
На первый взгляд тут, в адренергических и нейропептидных системах, есть обширнейшее поле для поисков интересующей нас разницы. Но тасуя теории, гипотезы, взгляды и результаты исследований по данному вопросу, мы обречены упереться лбом в простую догму, многократно подтверждённую экспериментально и возведённую в степень научного факта: «Сигналы в нейронах высоко стереотипны и одинаковы для всех животных» (Nicholls J. G., Martin A. R., Wallace В. G., Fuchs Р. A From Neuron to Brain, 2006).
Круг замкнулся. Вновь мы приходим к не слишком оригинальной мысли, что особые возможности позднего homo, т.е. интеллект, надо искать не в головном мозге, а вне его.
Думаю, даже не следует уточнять, что все те образы, слова, цифры и понятия, которые Эйнштейн так эффектно сложил в формулировки законов Вселенной, порождены не им самим, а теми поколениями, что последовательно, по крупицам созидали интеллект (фиксируя результаты создания на искусственных носителях), являющийся теперь общей собственностью homo.
Естественно, речь не только о прямых предшественниках Эйнштейна: Галилее, Ньютоне, Максвелле и Лоренце.
В такой же степени это относится и к шумерским жрецам, отследившим разницу скорости тела и его тени; к дикарю неолита, впервые зафиксировавшему символ звука своей речи с помощью странной царапины на стене пещеры; к крестоносцу, засмотревшемуся на обгорелый кусок бумаги с цифрами, начертанными по-арабски, и так далее. (Всех, кто сделал свой вклад в дело формирования коллективного интеллекта перечислить, разумеется, немыслимо.)
Мозг Эйнштейна лишь сконструировал из этих (и всех остальных) готовых компонентов новую модель понимания мира, оказавшуюся пригодной для того, чтобы стать ещё одним фрагментом общего интеллекта homo, и в свою очередь послужить деталью для будущих интеллектуальных конструкций.
Хотя всё могло быть и совершенно по-другому.
Предположим, маленького Альберта, в 1879 году, украли бы (к примеру) цыгане, во множестве кочевавшие по Швабии.
Предположим, эти цыгане, испугавшись полицейского досмотра, бросили бы малыша в (тогда диких) лесах Южной Германии, предоставив нашему гению возможность прожить лет пятнадцать в обществе лишь сов, бурундуков, вепрей и лисиц.
В результате мир обогатился бы ещё одним безымянным мауглеоидом. (Это мы знаем, что родители назвали его Альбертом, для него самого его же собственное имя осталось бы неизвестным).
Годам к шести Альберт Эйнштейн, вероятно, умел бы неплохо мычать и рычать.
Крайне сомнительно, что он овладел бы техникой швыряния камней, но подросши, он вполне мог бы практиковать изнасилования одиноких велосипедисток.
Возможно, со временем (в результате большой полицейской облавы) его чучело, с табличкой «Tarsanus Bavaricus» украсило бы музей естественной истории (к примеру) в Висбадене.
Конечно, и это тоже было бы (в известном смысле) вкладом Эйнштейна в науку, но с теорией относительности пришлось бы немного подождать.
До рождения следующего гения, которому ничто не помешает обучиться речи, алфавиту, счёту и нескольким тысячам слов и понятий.
На этом простом примере опять хорошо видно, что любые интеллектуальные действия возможны лишь при условии заимствования всех компонентов этих действий извне, из того коллективного интеллекта, о котором я уже говорил выше. Сам же мозг, каким бы он ни был, этих компонентов не содержит и не порождает.
Впрочем, в этом нет ничего нового или неожиданного.
Ещё Н. Harlow и М. Harlow в «Learning to Think» (Sci. Amer., 1949. Vol. 181) вывели точную, но корректную формулировку: «Мозг необходим для мышления, но необученный мозг недостаточен для осуществления этой функции, как бы хорош сам по себе он ни был».
J. Delgado в «Physical Control of the Mind» (1969): «Мозг человека, наряду с другими качествами, способен к изучению языков, к абстрактному мышлению и к моральным суждениям, но он не создаёт всего этого».
Ещё определеннее выразился в 1966 году С. Geertz, говоря о людях, не получивших по разным причинам доступа к «коллективному интеллекту»: «Они не будут, как это утверждает теория антропологии, напоминать одарённых обезьян, не нашедших своего места в жизни. Это будут чудовища с весьма ограниченным числом полезных инстинктов, ещё меньшим — чувств и полным отсутствием интеллекта» (Geertz С. The Impact of the Concept of Culture on the Concept of Man//Bull. Anatom. Sci., 1966. Vol. 22).
(Иными словами, единственной чертой, сегодня отличающей мозг homo от мозга других животных, является его успешность в деле обмена информацией внутри самого вида homo. Следует напомнить, что эта успешность целиком зависит от наличия общих систем номинации, созданных, вероятно, всего лишь 12-20 тысяч лет назад. Обмен информацией с представителями любого другого вида животных для homo либо невозможен, либо крайне ограничен самыми примитивными позициями. Достойно упоминания, что академическая нейрофизиология, в результате многих тысяч опытов, экспериментов, наблюдений и исследований — всего лишь подтвердила вывод Жюльена Офре де Ламетри, сделанный им ещё в 1745 году, в трактате «Естественная история души»: «Человек, до изобретения слов и знания языков — являлся лишь животным особого вида».)