Удаление второй группы («служебной») — действие чуть более спорное, но допускаемое эволюционной логикой, согласно которой анализаторы зарождались и образовывались лишь потому, что были востребованы сверхдревними структурами как инструмент оценки усложняющейся реальности.
К слову, даже очень далекий от всякого «центрэнцефализма» И. Павлов в своих итоговых трудах вынужден был признавать, что «подкорковые центры в большей или меньшей мере определяют деятельное состояние больших полушарий и тем разнообразно изменяют отношение организма к окружающей среде» (Павлов И. П. Двадцатилетний опыт объективного изучения высшей нервной деятельности, 1938).
Дальнейшие «удаления» уже рискованны, но они и не требуются. Вполне достаточно того, что имеющиеся факты предоставляют нам возможность означить целые структуры и отдельные формации нервной системы как «управляемые», а вероятно, и «подчинённые».
Исходя из вышесказанного, и у нас (вслед за Пенфилдом) появляется, наконец, известное право предполагать иерархичность всей нервной системы; a, следовательно (согласно самому принципу иерархии) — и наличие той самой «высшей точки», которая осуществляет интеграцию самой нервной системы точно так же, как нервная система осуществляет управление всем организмом.
Иными словами, спокойный и беспристрастный анализ структуры нервной системы, проведённый по принципу простого «исключения» формаций, вновь выводит нас на утверждение У. Г. Пенфилда о том, что «совершенно очевидно, что мозг должен иметь центральный координирующий и интегрирующий механизмы» (Пенфилд У. Г. Речь и мозговые механизмы, 1959).
Следует понимать, что идеально точно обозначить местонахождение этого «механизма», вероятно, пока невозможно. Но мы вполне можем довольствоваться и теми примерными координатами (ретикулярная формация ствола мозга), что обозначены творцами теории и подтверждаются логикой эволюционного процесса.
Следует отметить, что поиск интегрирующей «центральной системы» мозга отнюдь не замыкается в пределах «теории Пенфилда» или формулировок Ч. С. Шеррингтона или Г. Мэгуна. В разное время и с разной степенью опасливости к данной материи подступались выдающиеся нейрофизиологи и нейробиологи, даже те, что весьма прохладно относились к центрэнцефалическим гипотезам. В данном случае я говорю о Г. Шеперде, Д. Харрисе и (в какой-то степени) о Л. Рено.
Г. Шеперд хотя и призывал к «здоровому скептицизму» в вопросах поиска «нервного субстрата различных форм поведения» и констатировал, что «история исследований в этой области полна неудач и разочарований», тем не менее, на страницах своего итогового академического труда формулировал с предельной отчётливостью, что «центральные системы можно определить как совокупность клеток и нейронных сетей, выполняющих функции, необходимые для координированного поведения организма в целом. Как следует из этого определения, центральные системы не взаимодействуют непосредственно с окружающей средой. Но хотя они не являются составной частью сенсорных или двигательных систем, некоторые элементы последних могут относиться также и к центральным системам» (Shepherd G. Neurobiology, 1983).
Даже в этой краткой цитате легко заметить понимание Г. Шепердом неизбежности наличия центральной интегрирующей структуры мозга (у Г. Шеперда — «структур»). Специфичность школы, к которой принадлежал Г. Шеперд (а он был не нейрофизиологом, а нейробиологом), провоцировала поиск этой «центральной структуры» не в функциях древнейших формаций мозга, а исключительно в действиях нейроэндокринных механизмов этих формаций.
Свои взгляды Г. Шеперд базировал на исследовании аденогипофиза Дж. Харриса «Нервная регуляция гипофиза» (1955), на работах по нейроэндокринным клеткам гипоталамуса Л. Рено (Renaud L., 1977) и на изысканиях X. Костерлица и Р. Хьюза (1975) в области эндорфинов и энкефалинов.
Стоит отметить, что «нейроэндокринная версия» как гипотеза так и не состоялась. Экспериментальные, да и теоретические изыскания не позволили (в результате) хоть как-то соотнести наличие «центральных систем» с ролью эндорфинов, энкефалинов, гамма-аминомасляной кислоты, серотонина и так далее.
(При этом, естественно, остаётся несомненной важность влияния серотонинергической, норадренергической и других нейроэндокринных систем на общую работу мозга.)
Этот пример я привёл исключительно в качестве иллюстрации того, что вне зависимости от принадлежности к той или иной школе нейрофизиологии при понимании основных принципов работы и истории развития мозга поиск интегрирующей его силы неизбежен.
Для нас, в нашем исследовании, не является принципиально важным, где именно (или в чём именно), в структурах, процессах, пептидах, гормонах, кислотах и так далее, концентрируется и «обитает» это интегрирующее начало. Позволим себе ещё более «еретическое» утверждение: даже если будущие открытия «переместят» центрэнцефалическое начало из ретикулярной формации, к примеру, в базальные ганглии или в механизм влияния глицина, по сути исследуемого нами вопроса ничего не изменится.
Для нас важен лишь момент признания наличия такого начала, а уж с дислокацией и принципом проявления пусть разбираются будущие поколения нейрофизиологов, если сомнения в верности ретикулярной теории станут обоснованными и обретут доказательную базу. (Тема нашего исследования позволяет работать в первую очередь с принципами, а не только с терминами и механизмами.)
Итак, пока для такого «перемещения» нет никаких оснований, мы оставляем в качестве основного инструмента теорию Пенфилда и пользуемся принятой в ней терминологией.
Если из мира терминов нейрофизиологии вновь переместиться в область приблизительных понятий, то именно к этому «центральному координирующему и интегрирующему механизму мозга» вполне может быть применимо несколько размытое понятие «личность».
(Я употребляю его исключительно для того, чтобы не обращаться постоянно к прекрасной, но очень яркой формулировке Шеррингтона — «биологическая индивидуальность».)
В прошлой главе «личность» рассматривалась нами исключительно как неизбежная потребность и способность биологического существа самоидентифицироваться и выстроить своё поведение в мире в соответствии с этой самоидентификацией.
Сейчас, вероятно, придётся согласиться с тем, что её реальные функции значительно объёмнее и шире, чем простое «самосознание», что «persona» — это функция столь древняя, всевластная, глобальная и первоочередная, что всё происходящее в головном мозге свершается для неё и во имя неё, а она использует структуры мозга как инструменты, чтобы определять судьбу организма и его роль.
Тут, конечно, настораживает почти неуловимый, но всё же присутствующий «душок сверхъестественности». Впрочем, «душок» легко «отбить», просто признав размытое понятие «persona» литературнобытовым псевдонимом нейрофизиологического термина — formatio reticularis. Данная гипотеза может считаться естественным логическим продолжением всех представлений о ретикулярной формации, но она не согласуется с теорией А. Уорда, «разместившего» «личность» в 24-ом поле, а это уже лимбическая система (Ward A. The Cingular Gyrus: Area 24, 1948), и с представлениями Папеца, по которым «личность и эмоции» локализованы в третьем желудочке (Papez J. A Proposed Mechanism of Emotion, 1937). Это противоречие не может считаться решающим фактором, но упоминания, без сомнения, достойно.
Этот вывод создаёт иллюзию «открытия огромных перспектив постижения природы человека через эволюционную историю мозга», но, увы, это перспективы, преимущественно, для догадок, мало уместных в данном исследовании. Более того, «природа homo », не имеющая никаких принципиальных отличий от природы любого другого животного, никак не может рассчитывать на особое место в этой истории.
Единственное, о чём можно говорить с уверенностью, так это о том, что «управление» демонстрирует жесткую зависимость от данных сознания и результатов «адресации» к гиппокампальной базе; так как вся последовательность поведенческих актов, автором которых, вероятно, является т.н. личность, глубоко и прочно адаптирована к той реальности, в которой она находится в настоящую минуту.
Глубокая архаичность «личности», её возраст, насчитывающий как минимум 500 миллионов лет, сегодня порождает отнюдь не носоглоточный рёв и щелканье челюстями, а удивительную точность адаптации, адекватность предложенным условиям среды и правилам всевозможных межличностных или социальных игр.
(Как мы знаем, «игры» «живут» только в очень узком диапазоне особых искусственных обстоятельств, а при их разрушении или повреждении — изменяются, нарушаются или вообще аннулируются. Важным фактором является и постоянная потребность всякой «личности» в нарушении правил этих игр. Потребность обусловлена перманентным противоречием правил и агрессий, но это отдельный разговор, опять-таки находящийся вне темы данного исследования.)
Примечательна равнозначность, с которой «личность» на протяжении этих 500 миллионов лет моделирует любое адекватное поведение, без различий его отношения к «добру» или «злу», тем самым категорично выводя эти категории за пределы хоть сколько-нибудь существенных факторов развития жизни.
Было бы нелогично предположить, что сама сущность биологической «личности» могла претерпеть какие-либо серьёзные изменения со времён палеозоя, когда началось отчётливое и массовое оформление организмов в «сложноповеденческие» виды; когда все существенные факторы уже «вложились» в её формирование и, по всей вероятности, его завершили.
Поясняю.
Под существенными факторами следует понимать происшедшее в кембрийский период (545-490 млн. лет назад) появление праобраза зубов и зарождение обоняния, зрения и слуха.
Ордовик (490-434 млн. лет назад) наделил организмы феерическими возможностями, которые дают челюсти, т.е. ротовым аппаратом «хватательного типа».
Чуть позже (434-408 млн. лет назад) силур — добавил к челюстям глобальное удобство поперечноротости. Как справедливо отметил академик А. Северцов: «Новообразовавшиеся челюсти оказались органом настолько стойким, что несмотря на все другие крупные морфологические изменения, происшедшие в передней части головы потомков протогнатостом, они остались в общих чертах неизменными даже у наиболее поздних потомков этих древних форм. Действительно, ротовой аппарат хватательного типа, состоящий из верхней и нижней челюстей, одинаково сохранился как у рыб, так и у амфибий, как у рептилий и птиц, так и у млекопитающих» (Северцов А. Главные направления эволюционного процесса, 1967).
В силурийском же периоде возникают парные плавники и обретается тонко ориентированная скорость перемещения.
Пермский период (298-251 млн. лет назад) приносит смену сред (выход на сушу), обретение ног и локомоторные возможности передвижения в совершенно иной, по сравнению с «водной», реальностью.
Юрский период (205-141 млн. лет назад) даёт крылья, а меловой (141-65 млн. лет назад) — невероятные возможности плацентарности и млекопитания.
Обнаружить что-либо равное по значению этим факторам в дальнейшей истории «личности» позвоночных практически невозможно.
Речь и мышление homo были, без сомнения, любопытными новациями, но к числу существенных факторов причислены, всё же, быть никак не могут, так как не являются наследуемыми свойствами, а традиция их искусственной передачи может быть прервана любым катаклизмом, в отличие, например, от зрения или поперечноротости.
Принцип функции «биологической личности», по всей видимости, неизменен. Развитость этой «личности» может варьироваться (как варьируется, например, сила обоняния, острота зрения или чувствительность других рецепторов), но принцип остается неизменным, как остаются неизменными передающиеся от вида к виду генерирующие её стволовые структуры мозга.)
CAPUT XI
Эдиакарский период. Рай. Развитие нервной системы.
Разрушение рая. Явление протогерцины.
Основной инструментарий личности.
Война всех против всех.
«Проклятый» вопрос, который
благоразумно «обходят» все.
Поперечноротость богов. «Нумизматика».
Формирование мозга. Протокраниаты.
Гёте. Мнение А. Северцова.
Разумеется, реальный возраст т.н. личности, или биологической индивидуальности, вероятно, восходит к протерозою, к самым первым многоклеточным формам жизни в земной истории.
Dickinsonia costata, Cyclomedusa, Spriggina и другие (примерно 200) известные науке организмы эдиакарского (вендского) периода, судя по всем особенностям их морфологии, были лишены всяких агрессий (илл. 47).
У них не было возможности не только убивать, но даже и просто мешать жить любому другому существу. Слепые и глухие, не знающие ярости и амбиций, органокомплексы пластинчатого типа тихо подъедали донные отложения, без энтузиазма плодились и без всяких лишних переживаний заканчивали свои дни.

Илл. 47. Типичный вендобионт (обитатель эдиакарской эпохи) —
Dickinsonia costata
По всей вероятности, межнейронный обмен у вендобионтов осуществлялся преимущественно по простейшему «контактному» (асинаптическому) типу, наподобие того, что был обнаружен у известковых и кремниевых губок исследователями О. Тюзе (1953), М. Паванн де Секкати (1955), Г. Паркером (1910), Г. Мак-Ниром (1923); а сама нервная система, естественно, была диффузна и «редкорассеянна». (McNair G. Motor Reactions of the Fresh-water Sponge, 1923; Parker G. The Reactions of Sponges with a Consideration of the Origin of the Nervous System, 1910; Pavans de Ceccatty M. Le système nerveux des éponges calcaires et siliceuses, 1955; Tuzet O., Pavans de Ceccatty M. Les cellules nerveuses de l'éponge, 1953). Предположительно, что эдиакарский «рай» был обязан своей стагнацией ещё и тем, что он, возможно, частично воспринял от более ранних эпох (криогения, тония, стения, эктазия и так далее) то первоначальное положение, при котором «ещё не существовало никакой чёткой системы наследственности, никаких генетических механизмов, которые обеспечивали бы непрерывное воспроизведение функциональных единиц» (Ehrlich Р., Holm R. The Process of Evolution, 1963). Это предположение крайне дискутивно, но следует отметить, что и вообще все касающееся столь отдалённых от нас периодов и не может быть иным.
В этой протерозойской благостности ничто (судя по всему) не имело никакого значения: ни дивергенция, ни отбор, ни прогрессия возрастания численности, ни изменчивость. В течение примерно 100 млн. лет, уже нося на себе настоящую жизнь, юная Земля не знала ни страхов, ни борьбы, ни наслаждений.
Состояние интересующей нас функции (биологической индивидуальности) определить на данном этапе достаточно сложно; даже имея свидетельства наличия самой нервной системы, мы можем лишь гадать о её характеристиках.
С определённой уверенностью мы можем говорить лишь о полном отсутствии структур и рецепторов, способных формировать полноценное сознание и агрессии, а также о том, что толщина нервных волокон кротких вендобионтов была сопоставима с волокнами Aurelia, Metridium или Renilla, следовательно, колебалась от 2 до 12 мк. Соответственно, скорость прохождения импульса по таким волокнам не превышала 0,13-0,078 м/с, что категорически недостаточно даже для простой адекватной рефлекторики.
Существует строгое правило, в соответствии с которым о «скорости прохождения импульса» можно говорить только соотносительно с температурой тела, в котором происходят нервные процессы, или окружающей среды, если есть возможность обоснованного предположения, что тело принимает температуру среды. Применительно к эдиакарской эпохе это правило также должно соблюдаться, но климатические данные остаются пока весьма приблизительными, а принципы терморегуляции самих первосуществ и вовсе неизвестны. Единственное, что отчасти ясно, так это то, что за период с 580 до 550 млн. лет назад глобальных оледенений не наблюдалось, соответственно, возможно предположение, что вендобионты принимали температуру среды, которая была достаточно высока (температура эдиакарского моря около 40 °C). Такие температуры, теоретически, оптимальны для скорости прохождения нервных импульсов.
Поворотным пунктом развития жизни (временем разрушения эдиакарской идиллии) становится ранний кембрий.
Примерно 550 млн. лет назад эволюция свершила несколько циклов трансформаций, сперва обогатив (разветвив, укрупнив, утолстив) нервные ганглии, а затем заложив основу нейропиля (аналога ретикулярной формации у беспозвоночных).
Ветвление ганглий, обогатив чувствительность внутренних органов и поверхностей, «познакомило» организмы с множеством новых воздействий и раздражений. Эти раздражения (внешние и внутренние) потребовали верных «ответов» организма — и появились уже полноценные рефлексы, а как средство их оптимального «обеспечения» — синаптические связи меж нейронами. Мобильно объединяясь (по обстоятельствам) в специализированные «группы», рефлексы суммировались в агрессии, управление которыми стало возможно через зарождающийся нейропиль. Толщина нервных волокон увеличилась, а с ней вместе возросла и скорость прохождения импульсов.
Теоретически можно предположить, что к самому концу протерозоя диаметр нервного волокна был сопоставим с тем, каким располагает, к примеру, полихета Neanthes (30-37 мк), и соответственно, скоростью проведения импульса около 5 м/с; а к началу кембрия его толщина возросла до 70-150 мк, а скорость составила 10-15 м/с, как у Cambarus. (Приводится по: Prosser L., Brown F. Comparative Animal Physiology, 1962. Табл. 54.)
По свидетельству X. Коштоянц (1957), Т. Бэллока (1943), именно у кишечнополостных, разрушивших усложнением своей НС эдиакарский «рай», уже возникает синаптическая, сложная и богатая связь между нейронами (Bullock Т. Neuromuscular Facilitation in Scyphomedusae, 1943; Коштоянц X. Основы сравнительной физиологии, 1957. Т. 2).
Прогрессирующая чувствительность рецепторов и их количественное умножение — обострили половое чувство, голод, страх и боль. Первые, ещё крайне наивные попытки соперничества за самок и еду — породили намёк на рефлекторику ярости, конфликты и далёкое предощущение вкуса чужой плоти.
«Личность», с её агрессиями, потребностью самоутверждения, конфликтностью, чувственностью и способностью чёткой самоидентификации буквально «сплеталась» из густеющей сетки нервных волокон.
Впрочем, пока это был лишь её тусклый эскиз, так как «на самой низшей ступени животного царства чувствительность является равномерно разлитой по всему телу, без всяких признаков расчленения и обособления в органы. В своей исходной форме она едва ли чем отличается от так называемой раздражительности некоторых тканей, например, мышечной у высших животных...» (Сеченов И. М. Элементы мысли, 1952).
Конец эдиакарской эпохи, начало кембрия — это то время, когда «личность» уже обозначилась как один из важнейших инструментов эволюционного процесса и (возможно) его генеральный стимулятор. Она ещё инкубировалась, но развивающаяся и крупнеющая нервная система уже переполняла древние организмы ищущей выхода и реализации рефлекторикой.
Именно «личности», её главным свойствам (т.е. потребности в нападении, защите, в реализации через конфликт), и предстояло, взорвав эдиакарский рай, сдвинуть жизнь с «мертвой точки», на которой та пребывала 100 миллионов лет. Но для того чтобы это произошло, наша «биологическая индивидуальность» нуждалась хотя бы в минимальном инструментарии.
Этот инструментарий «зрел» сравнительно недолго, и уже в фортунском «веке» раннего кембрия, а это (примерно) 545 миллионов лет назад, на сцене эволюционного театра являются первые протоконодонты, такие как, например, малышка (около 2 мм) Protohertzina. У неё ещё не было ни глаз, ни челюстей — лишь скопление зубоподобных шипиков, но этого оказалось достаточно: вскоре кембрийские воды впервые познали убийство, и большой механизм животной эволюции благополучно запустился.
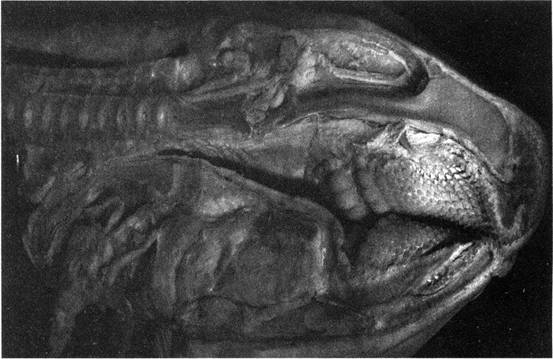
Илл. 48. Дентальный аппарат акулы
Крохотуля Protohertzina по праву может считаться условной точкой отсчёта в истории «личности»: впервые живое существо, исходя из своих собственных побуждений и потребностей, начинает влиять на среду и провоцировать её множественные изменения.
«Шипики протогерцины» пережили сотни эволюционных метаморфоз; они стали клыками, молярами, резцами, жалами, когтями, рогами, клювами, педицелляриями и бивнями. Но переданные даже самым далеким потомкам и радикально видоизменённые, «шипики» не утратили своего первоначального предназначения, и вот уже 545 миллионов лет являются основным инструментом «личности». (Клинки, шпаги, штыки, стилеты, да и вообще всё оружие homo — это ведь тоже не более чем очередное перерождение «шипиков» раннего палеозоя).
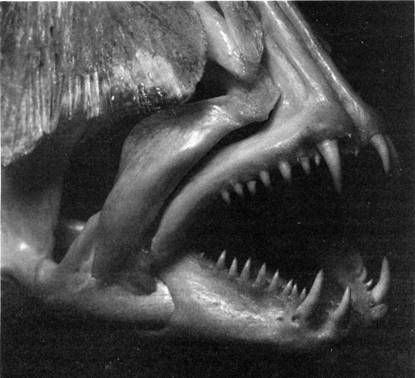
Илл. 49. Дентальный аппарат костной рыбы
Развитие и многоипостасность «шипиков» спровоцировали и развитие эффективных защит от данного «инструмента личности». Возникли: раковины, простые и склерозированные кутикулы, чешуя, кожа, роговые щитки, шерсть, хитиновые карапаксы, мимикрия, скорость локомоции и так далее, благодаря чему разнообразились формы активной жизни. В свою очередь «совершенствование защит» спровоцировало организмы на генерацию более совершенного «оружия» и так далее и до бесконечности.
Цена развития оказалась, впрочем, не очень высокой. Со времён протерозоя было уничтожено без всякого следа примерно 99,9% всех видов живых существ, что не так уж много, учитывая, что именно конфликтность, постоянная война всех против всех являлась и является обязательным условием и движителем эволюции. (Разумеется, не все эти виды были истреблены конкурентами, существенная часть погибла в катаклизмах, переродилась или была «списана», по разным причинам, самим эволюционным процессом.)
Здесь становится ещё более понятна природа и назначение той функции головного мозга, которую мы ранее обозначили как биологическую индивидуальность («личность») и за которой условно закрепили «потребность и способность в точной и постоянной самоидентификации организма».
Но наш краткий экскурс в эдиакарскую и кембрийскую эпохи позволяет обоснованно предположить, что у «личности» есть (как минимум) и ещё одна задача, по важности сопоставимая с самоидентификационной — способность управлять рефлексами и концентрировать их на том или ином обстоятельстве, противопоставляя этому обстоятельству сумму управляемых рефлексов, т.е. агрессию.
В переводе на язык приблизительных понятий я бы обозначил это свойство одновременно и как генерацию конфликтности, и как её строжайшую регуляцию.
Напомню: когда-то именно конфликтность была тем важнейшим фактором, которого недоставало эдиакарской эпохе, чтобы от стагнации перейти к развитию; но как только организмы получили возможность конфликтовать друг с другом и средой — выход из тупика был найден, и настоящая жизнь началась.
Вглядимся и оценим: нейрон по самой своей природе исключительно конфликтен, грубо говоря, конфликтность (т.е. обеспечение реакции организма на обстоятельство) и есть его основная биологическая специализация.
С учётом того, что большинство «обстоятельств», предлагаемых средой — неблагоприятно или прямо враждебно, конфликтная роль нейрона становится ещё более понятна.
Разумеется, объединенные в сложные сети и цепи, связанные меж собой в многомиллиардные сообщества (как дифференцированные, так и сегрегированные), нейроны способны к исключительно мощной и скоростной конфликтности, которая мгновенно «воспламеняет» весь организм, творя поведенческий ответ среде или обстоятельству.
Но мы знаем, что нервная деятельность состоит не только из возбуждений, механизм которых более или менее понятен, но и из торможений, когда вся сила нейронального «ответа» на обстоятельство может быть употреблена либо на погашение (ослабление или недопущение) рефлекса, либо вообще на изобретение парадоксальных (соотносительно с обстоятельствами) форм поведения.
Здесь следует признать, что происхождение и природа такого существенного фактора, как торможение нервной системы, так и осталась, мягко говоря, крайне дискутивной.
Конечно, ситуация становится ясна, когда мы, подобно Ивану Михайловичу Сеченову, посредством малюсенького пинцетика, кладем на обнажённый ствол мозга лягушки — кристаллик соли и видим торможение почти всех моторных процессов, угнетение части рефлексов.
«Этот эффект развивается обыкновенно в течение первой минуты по приложению к мозгу раздражающего вещества, прежде чем успеет раздражение вызвать какое-либо движение (прямое или отражённое). Бывают, однако, случаи, когда угнетение отражённой деятельности заставляет ждать себя дольше. Тогда лучше всего вместо раствора приложить к мозгу кристаллы соли, которые действуют энергичнее и скорее... эффект раздражения мозга солью постепенно исчезает, если раздражитель удален (несколько капель воды на мозг и обсушивание его пропускной бумагой)» (Сеченов И. М. Исследование центров, задерживающих отражённые движения в мозгу лягушки, 1863).
Конечно, «соляные опыты» Ивана Михайловича лишь продемонстрировали факт искусственно вызванной временной травмации нервной системы и сопутствующих ему состояний организма, т.е. угнетения.
(Термин «торможение», когда-то предложенный ещё Декартом, будет введён в нейрофизиологический обиход самим же И. М. Сеченовым, но чуть позже. Выше мы процитировали самую первую из девяти его «лягушачьих» работ.)
На первый взгляд ничего удивительного в этих опытах не было, так как любая травма, особенно важных структур, вызывает «угнетение» организма.
Примечательным в этом простом опыте было лишь то, что возникшие при искусственной травмации состояния были аналогичны тем, что возникают и без всяких «кристалликов соли», т. е. благодаря лишь процессам, естественно происходящим в механизмах мозга.
Тут-то и стало понятно, что совершено открытие не просто сверхважного механизма, а возможно, коренного принципа работы нервной системы.
Конечно, сами по себе опыты были очень грубым выявлением этого принципа, а результат каждого из них — предельно условной моделью реально происходящего в головном и спинном мозге.
К слову, естественно, И. M. Сеченов использовал не только соль, но и другие травматоры: кислоты и электричество. Средства воздействия намеренно были избраны столь жёсткие, с таким мощным некротизирующим потенциалом. Дело в том, что И. М. Сеченов и не ставил своей целью выявить агент, провоцирующий торможение, важным было лишь наличие самого его факта, его продолжительность, нюансировки, влияния.
Итогом «лягушачьих» экспериментов Сеченова, продолжавшихся почти пять лет и подробно описанных в девяти его научных работах45 стало понимание факта, что нервная система, управляемая только возбуждающей силой, нежизнеспособна и обладает тенденцией к «саморазнесению», к уничтожению организма, в котором живет.
Только при наличии молниеносного контроля за силой и нарастанием рефлекса (за потенциальной конфликтностью нейрона) нервная система способна создать вокруг себя управляемый организм и «провести» его сквозь внешнюю среду, обеспечив ему нормальную биологическую этапность: рождение-размножение-гибель.
Стоит отметить, что формальным первооткрывателем явления был (как я упомянул выше) Рене Декарт (Картезий), попытавшийся разъяснить движение глаз «возбуждением» и «расслаблением» соответствующих глазных мышц. В 1854 году братья Веберы, раздражая блуждающий нерв, сумели понизить частоту сердечных сокращений, что добросовестно описали, но должного значения своему открытию не придали, хотя их эксперимент был прямым доказательством именно тормозящих факторов (Weber Е. F., Weber Е. Н. Experimenta, quibus probatur nervos vagos rotatione machinae galvanomagneticae irritates, motum cordis retardare et adeo intercipere, 1845).
Глобальность роли «торможения» несомненна, обсуждать её даже несколько странно; причём не только по части контроля над рефлексами, но и в деле их формирования. Понятно, что при «вычитании» торможения становятся невозможными не только все сложные физиологические процессы и все поведенческие акты, но даже и такой банальный вид рефлексов, который именуется «тоническими».
К примеру: «Тонические рефлексы относительно медленные, длительно продолжающиеся установки, которые поддерживают мышечный тонус, позу и равновесие» (Dethier V., Stellar Е. Animal Behaviour: Its Evolutionary and Neurological Basis, 1967). Понятно, что эти, да и ещё множество других рефлексов и проявлений — в чистом виде «дети» торможения, а не возбуждения. Таких примеров можно было бы привести множество, но все они давно являются общим местом, следовательно, общеизвестны.
С того момента, когда пинцет И. М. Сеченова впервые возложил кристаллик поваренной соли на обнажённый лягушачий мозг, прошло почти 150 лет, но физиологический принцип торможения остался, по выражению И. П. Павлова, «проклятым вопросом» (Павловские среды, 1949. Т. 2).