Чтобы понять причину возможностей мозга homo оперировать номинациями, понятиями и образами, порождёнными вне его, необходимо разобраться с тем, что можно было бы назвать биологическим «прожиточным минимумом» этого мозга, т.е. с его врождённым, базовым инструментарием оценки реальности.
Наличие этого «прожиточного минимума» несомненно; его отсутствие не позволило бы виду выжить, а речь, мышление и интеллект, будучи явлениями недавними и искусственными, могли бы «обосноваться» лишь на свойстве более простом, получаемом всяким живым существом комплектно с субстратом мозга.
Здесь мы впервые подошли к понятию «разум», или же «ratio».
К слову, понятно, что это наименование достаточно условно, как, впрочем, и всякое другое слово в современном лексиконе, обозначающее столь неявные и малоизученные процессы.
Именно ему можно формально делегировать функцию этого простейшего, базового свойства, занимающего в иерархии всех оценочно-ассоциативных процессов (формально) самую низшую ступень. («Низшесть» ступени — определение, разумеется, дискутивное, порождённое скорее стереотипом, чем реальным знанием и положением вещей.)
Чтобы точно и наглядно проиллюстрировать данную тему, представим себе некую позвоночную «форму жизни», продвигающуюся по первобытному лесу.
Это может быть практически любой организм: особенности ног, ноздрей, мышечной массы, черепа или глаз — в данном случае малосущественны. Его цели тоже не играют особой роли; это может быть поиск плодов или полового партнера, преследование жертвы или, напротив, попытка спасения.
В любом случае, мозг этого существа проделывает некую оценочно-ассоциативную работу, без которой ни само движение, ни выполнение любой, даже простейшей задачи, невозможно.
Схема этой работы стандартна и применима к позвоночному любой геологической эпохи, от офиакодона (Ophiacodon) до мауглеоида; «показания» тактильных, термоцептивных, проприоцептивных, висцероцептивных, обонятельных, зрительных, слуховых и так далее рецепторов организма, неизбежно суммируются в «картинку» сознания. Нейроны гиппокампа фиксируют эту картинку, сохраняя обширную, точную, безначальную и бесконечную «ленту» подробностей мира, запечатленную суммой физиолого-рецепторных ощущений, т.е. сознанием.
Стоит отметить, что данные представления являются не моей новацией, а догмой нейрофизиологии. Ещё И. М. Сеченов писал: «Подобно фонографу Эдисона она записывает, сохраняет и воспроизводит внешние воздействия, но оставляет неизмеримо далеко за собой все чудеса этого инструмента. В самом деле, фонограф отвечает только на звуковые явления и записывает только данный индивидуальный случай. Память же вносит в свои реестры все вообще воздействия на все пять органов чувств (занося туда же все колебания мышечного чувства) и записывает не один данный ряд впечатлений, а миллионы их... Вот эта-то таинственная работа, начинающаяся в раннем детстве и длящаяся всю жизнь, и составляет то, что называют переработкой сырого впечатления в идейном направлении. И как ни сложна эта работа, как ни велика перетасовка составных частей впечатлений, тем не менее, память сохраняет запечатленные в ней образы и звуки настолько неизменно, насколько человек узнает образы и звуки как уже виденные и слышанные» (Сеченов И. Предметная мысль и действительность, 1908).
Ретикулярная формация ствола возбуждает и тормозит активность проекционных и ассоциативных зон коры, дирижируя как самими анализаторами коры, так (возможно) и их адресацией к «ленте гиппокампа», хранящей все ощущения, впечатления и наблюдения существа с момента его рождения.
Мы уже касались этой темы в главе VII, но здесь необходимо сделать ряд ремарок, поясняющих невозможность осуществления даже самых простых процессов разума без этой «адресации», и соответственно, без того, что И. М. Сеченов называл «актом узнавания».
Как я уже говорил, организм, мозг которого не способен сохранять классифицированные (в той или иной степени) впечатления (знания) и сопоставлять их с увиденным в настоящую минуту, будет ежеминутно обречён начинать познание мира заново, следовательно, не может быть и речи ни о каком адекватном поведении.
(К примеру, процесс изучения свойств, примет, запаха, вкуса и значения даже обычного куска скалы (не говоря о более сложных предметах, ситуациях, явлениях или формах жизни), теоретически, будет бесконечным и единственным занятием.)
Ещё одним (косвенным) подтверждением моих выводов о работе механизмов мгновенного «опознания образов» могут служить многочисленные наблюдения за различным лабораторным и клиническим материалом, страдающим одной из самых тяжёлых форм расстройства работы головного мозга, вероятно, вызванной неспособностью корковых центров «адресоваться» к гиппокампу.
Одним из первых эту патологию описал Н. Милк в своём труде «Über die Funktionen der Großhirnrinde» (1881), причём он сформулировал её всего в четырех словах: «Видит, но не узнает». Впрочем, ещё раньше Мунка Жан-Мартен Шарко (1847) объяснял это расстройство «утерей памяти на зрительные образы», а X. Джексон в 1876 г. фиксировал «нарушение зрительного восприятия (imperceptions), при котором больной не в состоянии узнавать предметы, которые он видит».
Эта дисфункция, в силу её драматических последствий и полной необъяснимости, долгое время казалась многовариантной, порождённой самыми невероятными процессами, и соответственно, имела множество имен и обозначений: «онероидное состояние сознания», «объектная агнозия», «симультанная агнозия Вольперта», «делириозное помрачение сознания», «оптико-пространственная агнозия», «зрительная асимболия», «апперцептивная агнозия Лисауэра» и так далее. Различные трактовки её происхождения предлагались Клейстом (1934), Ланге (1936), М. Кроль (1933), Балинтом (1909), Холмсом (1919), Вольпертом (1924), Ажуриагеррой и Экаэном (1960), Б. Биренбаумом (1948), Лурией (1959), Гельбом и Гольдштейном (1920) et cetera.
Здесь я перечислил лишь исследователей, которые (в указанные мною годы) занимались строго оптическими агнозиями. Основной причиной невозможности оптического узнавания предметов и образов считалась очаговая дефектация различных областей затылочно-теменной коры, но уже в шестидесятые годы XX века О. Успенской в двух трудах «Неврологическая клиника и диагностика опухолей боковых желудочков» (1951) и «Клиника опухолей III желудочка» (1959) были суммированы 156 наблюдений, которые, в известной степени, свидетельствовали о возможности возникновения различных агнозий вследствие патологий ventriculus lateralis или III желудочка. Сам по себе этот факт ничего не объяснил, но частично дезавуировал как «затылочно-теменную», так и другие «очаговые» теории.
Фонтанирование гипотезами (самыми невероятными) прекратилось лишь в последней четверти XX столетия; трактовки происхождения агнозий стали предельно скупыми и осторожными. Такие академисты, как Б. Зейгарник и Б. Биренбаум ограничились лишь описанием агнозических симптомов и публикацией протоколов исследований.
Процитирую, для примера, лишь малую часть описанных опытов: «Патопсихологическое исследование выявило грубое нарушение узнавания предметов. Больная часто не узнавала (40 %) предъявленных ей изображений. Так, нарисованный гриб она называет “стог сена”, спички — “кристаллами”». Выдержки из протоколов: 1. (Больной демонстрируют пионерский барабан. — Прим, автора); её оценка: «Горшок с кисточкой. Булка, которую кладут в кастрюлю, а это кренделёк (указывая на палочку). Похоже и на шапку». 2. (Экспериментатор показывает карандаш). Ответ: «Свеча, тут уж ясно, что свеча». 3. (Производится демонстрация изображения самолёта). Ответ: «Это стрелка. Это балкон, но причём тут стрелка и две ножки?» (Зейгарник Б. Патопсихология, 1976).
(К слову, нетрудно заметить, что во всех описаниях и протоколах (этих и других авторов) мы можем наблюдать только примеры относительно лёгкого поражения важнейшей функции «адресации». Уместно предположить, что формы более тяжелые вообще исключают всякий «контакт», делая экспериментальную работу невозможной.)
Разумеется, в столь серьёзном вопросе любой категоризм неуместен; в том числе и категоризм отрицания «затылочно-теменных» и прочих «очаговых» гипотез, которые, возможно, ещё будут «реабилитированы» получением новых экспериментальных данных. (В конце концов, если подтвердится любая из «очаговых» версий, это будет лишь свидетельствовать о нарушении правильности или корректности «запроса» ввиду повреждения «запрашивающих зон».)
Дело вообще совершенно в другом.
Существует неоспоримая очевидность того, что причиной невозможности «опознать» и, соответственно, «оценить» компоненты реальности (или реальность в целом) является некий нейрофизиологический «срыв» (механизм которого пока непонятен), сделавший для больного невозможным взаимодействие оперативных корковых структур восприятия с «базой» сознания.
(А вот произошёл ли «срыв» по причине невозможности корковых зон «адресоваться» или гиппокампальных — «ответить» на эту адресацию; или же причиной дисфункции стала третья причина, к примеру, патология проводящих путей меж этими отделами мозга, для данного исследования не является принципиальным вопросом.)
Строго говоря, на данный момент в оценке этого дефекта работы мозга нейрофизиология вернулась на уровень реплики Г. Мунка: «Видит, но не узнаёт».
А теперь вновь вернёмся к нашему «существу», его продвижению и его возможностям адекватно оценить реальность вокруг себя.
Мы не знаем, сколь на самом деле «груба» его адресация к «ленте» собственного сознания. Можно предположить, что отсутствие номинаций (т.е. развитого «языка»), т.е. полного и понятного классификатора многообразия мира, позволяет осуществить лишь самую незатейливую, а возможно, и не всегда успешную адресацию к той базе ассоциаций, что находится в нейронах гиппокампа.
Тем не менее, результаты даже этой примитивной адресации, позволяют точно и симфонично активировать нужные участки коры, отсекая второстепенные на данный момент факторы, а точная дирижировка агрессиями (через ретикуло-лимбические связи) — даёт возможность выстроить безошибочное и эффективное поведение.
Любопытно, но эта связка «грубой адресации и ответной ей аналогии или ассоциации» — самый вероятный претендент на роль того, что можно было бы назвать «разумом», тем самым прожиточным минимумом, который является врождённым и обязательным свойством мозга любого млекопитающего, а возможно, и «просто» позвоночного.
В любом случае, перефразируя известное высказывание Феодосия Добржанского (1900-1975): «Ничто в нейрофизиологии не может быть понято вне контекста эволюции».
Понятие «разум», несмотря на всю его кажущуюся расплывчатость, строго нейрофизиологично и эволюционно.
Оно привязано как к истории самого субстрата мозга (головного и спинного), так и к эволюционному усложнению физиологии организма.
Полагаю, здесь будет уместным уточнить, что исследования ассоциативных возможностей мозга млекопитающих и рыб начались ещё в 1927 году группой учеников И. Павлова Г. Зелёным, Г. Прокофьевым, В. Мапченковым. Уже в 1934 году в «Материалах V Всесоюзного физиологического съезда» была опубликована работа Н. Подкопаева «Условный рефлекс как ассоциация», в 1952-55 годах — труды Н. Рокотовой «Образование временных связей в коре головного мозга собак при действии нескольких индифферентных раздражителей» и «О временных связях на индифферентные раздражители у антропоидов (шимпанзе)», О. Малиновского «Выработка временной связи на индифферентные раздражители у кроликов» (1953), Н. Тих «Исследование процесса ассоциаций у детей и низших обезьян» (1955), И. Кармановой «Материалы к сравнительной физиологии коркового замыкания» (1954), Б. Сергеева «Эволюция ассоциативных временных связей» (1967), Ю. Трошихиной «Эволюция мнемической функции» (1973) et cetera. Вышеупомянутые работы фиксировали эксперименты по бесспорному выявлению реальных ассоциаций у высших и низших обезьян, кроликов, хорьков, морских свинок, собак, крыс, кошек, голубей, канареек, синиц и рыб.
Чуть выше я характеризовал процесс доречевой адресации как «весьма незатейливый» и даже «примитивный». Полагаю, имеет смысл попытаться оценить степень этой «примитивности». Именно для этой оценки нам и понадобится наше гипотетическое, дикое и крайне архаическое «существо».
Итак, что же происходит вокруг него, какова ежесекундная реальность «древнего леса», в котором оно продвигается?
Что именно должно быть оценено его мозгом с максимальной отчётливостью, чтобы «существо» оставалось (какое-то время) живым и сытым?
Какой набор точных ассоциаций, аналогий и контраналогий необходим для простого продвижения в мезозойских (к примеру) зарослях хотя бы на пять метров вперед?
Для начала разберёмся с тем, как выглядел этот лес и что он собой представлял.
Напомню, что в те времена, наряду с унаследованными от палеозоя папоротниками, хвощами, саговниками и гинкго, уже окончательно оформились хвойные. Ещё были «живы» весьма неторопливо уходящие с эволюционной сцены беннеттиты. Появились первые покрытосеменные и первые цветы, схожие с магнолиями и лотосами.
(По всей вероятности, уже существовали пигменты, изменяющие цвет листвы в зависимости от времени года. Но утверждать это с полной уверенностью трудно, так как девон, карбон и пермь, возможно, ещё не знали сезонного пигментирования.)
В общем, выбранный нами наугад период (поздний мезозой, т.е. «мел») — это шумная, яростная и яркая эпоха. Это время многотонных спинозавров и тиранозавридов; время яркоглазых малышей лиелиназавров и необычайно проворных бегунов — велоцирапторов; время флегматичных пожирателей мха — трицератопсов, да и ещё примерно тысячи видов, среди которых уже освоились такие биологические новации, как муравьи, сумчатые и плацентарные, а первая пернатая мелочь, типа иберомезорнисов (Iberomesornis), встретилась с уже «отходящими» птерозаврами, размах крыльев которых достигал 12-15 метров.
Залитый кровью, полный гулом, рёвом, драмами и нежностью (к детёнышам и половым партнёрам) поздний мезозой стал временем, когда формировались артикуляторы, герифицировались полушария, а на агрессии (в предчувствии скорого господства млекопитающих) наносилась самая последняя «полировка».
Исходя из вышесказанного вполне уместно предположение, что картина позднемезозойского леса была звучной и сочной.
Следовательно, для успешного движения по нему требовалось принципиальное «понимание» множества различных явлений, звуков, качеств, запахов и свойств. (Т.е. успешная адресация ассоциативных центров коры к гиппокампу, получение «ответа», позволяющего сопоставить находящееся перед глазами с тем, что хранится на «ленте» сознания. Заметим, что только это, в любую геологическую или историческую эпоху, позволяет сделать верную «оценку» реальности, а в зависимости от оценки, выстроить верную модель поведения).
Поясняю.
В «верхнем» поле зрения нашего гипотетического «существа» неизбежно находятся несколько тысяч разновеликих древесных листьев. Все они (соотносительно с размером и тяжестью) — в состоянии различной динамики. Соответственно, различны и межлиственные просветы, а через это — и образованный ими световой мерцательный узор, изменяющийся каждое мгновение.
Уже одна эта простая «картинка» требует наличия «поясняющих ассоциаций», которые позволят «существу» классифицировать множественность подвижных светлых и темных форм над своей головой как нечто малосущественное и почти не содержащее угрозы.
Причём стоит отметить, что излишняя «навязчивость» этих «пояснений» могла бы разбалансировать всю тонкую картину восприятия, сделав не самый важный фактор основным событием мгновения, что (опять-таки) могло спровоцировать неверное поведение. Как осуществляется «иерархия» значимости видимого? Гипотезу способности мозга отделить малосущественное предлагает в своём труде В. Д. Глезер: «Как показывают исследования зрительного восприятия, в модели мира, создаваемой мозгом, помимо механизма классификации участвует второй механизм... Функция второго механизма — определение пространственных соотношений; роль этого механизма самым тесным образом связана с актом внимания. По существу обе функции — конкретизация объекта за счёт описания пространственных отношений и внимание — представляют собой нераздельное целое» (Глезер В. Д. Зрение и мышление, 1985). Сущностная часть этой трактовки — в понятии «пространственных соотношений», т.е. в возможности дифференцировать «ближнее-среднеближнее-среднедальнее-дальнее», что подтверждается схемами движения глаз при рассматривании различных пейзажей, приведенными А. Ярбусом в его классическом труде «Роль движения глаз в процессе зрения» (1965). Понятно, что лес на картинах, послуживших материалом для экспериментов, совсем иной (чем в нашем случае), но принцип разной фиксации на разноудаленных деталях пейзажа — проиллюстрирован на схеме А. Ярбуса вполне убедительно (илл. 44а-b).


Илл. 44a-b. Схема движения глаз (на примере картины «Утро в сосновом лесу»). (По Ярбусу)
Чуть ниже уровнем (прямо перед глазами «существа») располагается многомерно-слоистая и тоже очень подвижная картина фронтального (уже не имеющего просветов) лесного фона, в котором перемешаны ветви, побеги, листва, лучи, стволы, стручки, тени, воздушные радикулы, плоды, цветки, стробилы, капающие смола и соки, льющийся птичий помет, осыпания пыльцы, падения отслоившихся пластин коры и пар, восходящий от горячих прикорневых болот. Этот фон динамируется срывами и пролетами птиц; перемещениями дейнозухов (Deinosuchus), протаскивающих свои двенадцатиметровые туши через прикорневые плетения; роющими мох эуплоцефалами и, возможно, ещё десятками представителей различных видов активной жизни, колеблющими и сотрясающими великолепное растительное месиво «мела».
В этой достаточно сложной динамической картине — значение имеет всё. Любой неверно понятый фактор может быть фатален. Не только зрительные, но и звуковые, и запаховые «гаммы» должны быть «узнаны» и молниеносно оценены по степени их опасности или нейтральности.
К примеру, резкое усиление запаха мха может означать его раздирание и жевание каким-нибудь тихим хазмозавром, т.е. может игнорироваться как малозначительный фактор. А вот аромат темпскии (Tempskia), внезапно подмешавшийся в общую запаховую «гамму», может означать чрезвычайную опасность, свидетельствуя о том, что бок дейнозуха только что резко «шаркнул» по основанию ствола этого папоротника (возможно, в броске), и надо спасаться, не особо вдаваясь в подробности, куда и на кого направлен бросок.
Разумеется, факторов, заслуживающих внимания «существа», больше в десятки раз, т. к. его мозгу необходимо принимать во внимание и точно оценивать состояние грунта под лапами, различные препятствия, сотни звуков, метания собственной тени и множество личных проприоцептивных и висцероцептивных показателей.
Значимость последнего фактора (несмотря на скупость моего упоминания о нём) не только несомненна, но по важности едва ли не равняется основным внешним раздражителям. Здесь уместно сослаться как на И. Павлова, так и на И. М. Сеченова: «Надо признать в больших полушариях существование ещё особых анализаторов, которые имеют целью различать огромный комплекс внутренних явлений, происходящих в самом организме. Нет сомнения, что для организма важен не только анализ не только внешнего мира, для него также необходимо сигнализирование вверх и анализирование того, что происходит в нём самом» (Павлов И., 1912); «Мышечное чувство может называться ближайшим регулятором движений и в то же время чувством, которое помогает животному познавать в каждый данный момент положение в пространстве, притом, как при покое его, так и при движении. Оно представляет, следовательно, одно из орудий ориентации животного в пространстве и во времени» (Сеченов И. М., 1891). (Добавлю, что любые висцероцептивные и проприоцептивные ощущения точно так же, как и любые внешние раздражители, нуждаются в «оценке» мозгом, т.е. в полноценном «акте узнавания».)
Не следует забывать и то, что наше «существо» тоже находится в движении, следовательно, постоянно получает как существенные изменения верхней, фронтальной и «подножной» картинок, так и смену «личных» ощущений.
Прошу отметить, что этот анализ может дать представление об оценочно-ассоциативной работе мозга лишь во время простого движения на дистанции в пять метров.
Разумеется, описанная мною фактура существенно уступает в сложности и многообразии любым «житейско-бытовым» или экстремальным ситуациям: ухаживанию-спариванию, драке, обучению детёнышей, обустройству логова, преследованию добычи, спасению от преследователя и так далее.
Полагаю, что говоря о работе «примитивного» врождённого разума, обеспечивающего элементарное выживание, мы говорим об обязательном «узнавании» и чёткой дифференциации примерно тысячи предметных позиций, о нескольких тысячах связей меж ними и о сложно поддающихся подсчёту количестве различных «понятных» явлений и (опять-таки) «знакомых» связей меж ними.
К слову, подобное явление заметили ещё И. Павлов и И. Сеченов: «Большими полушариями собаки постоянно производится в разнообразнейших степенях как анализирование, так и синтезирование падающих на них раздражителей, что можно и должно назвать элементарным, конкретным мышлением. Это мышление, таким образом, обусловливает совершенное приспособление, более тонкое уравновешивание организмом окружающей среды». (Павлов И. Двадцатилетний опыт объективного изучения высшей нервной деятельности (поведения) животных, 1952); «В узнавании есть... даже элементы рассудочности, настолько процесс напоминает собой умозаключительные акты» (Сеченов И. М. Элементы мысли, 1935).
Трудно предположить, что точная оценка столь значительного количества реалий может иметь какой-то другой механизм, кроме работы проекционных и ассоциативных зон коры, их успешной адресации к «базе сознания», хранящейся в гиппокампе, и безупречности связей как коры, так и «базы» с некой интегрирующей церебральной структурой, возможно, с ретикулярной формацией.
Здесь я обрисовал лишь ничтожно малую часть происходящего в головном мозге нашего архаичного «существа». Но даже эта малая часть даёт представление об основном принципе и симфоничности его работы.
Исходя из вышесказанного, простейший разум, который есть врождённое и неизбежное свойство любого мозга, «примитивным» назвать будет очень сложно. (Хотя пафосность этого понятия несколько и страдает при его трезвой нейрофизиологической оценке.)
Более того, вновь становится почти неизбежным вывод о том, что «мышление» не является никакой принципиальной новацией, что это лишь укрупнение, расширение и обогащение древнейшей функции мозга, происшедшее благодаря изобретению речи, т.е. номинативной системы.
Тут, разумеется, возникает множество вопросов.
И первый вопрос — почему именно homo оказался изобретателем и обладателем этого (возможно) выигрышного «лотерейного билета» (номинативной системы), а не другое млекопитающее животное? Этот вопрос, в известной степени, любопытен, но не является первоочередным. (Следует помнить о микроскопичности как самого фактора «homo», так и его интеллекта в истории развития мозга позвоночных.)
Гораздо важнее определиться с тем, а кому, собственно (в границах любого организма), «принадлежат» все эти ощущения, знания, агрессии и другие инструменты приспособления к реальности и влияния на неё, включая и простейший «базовый» разум?
Кто «хозяин» и «пользователь» всего этого?
«Кто» эта точка организма, концентрирующая в себе часть эффекта могучей электрохимии мозга и использующая её как руководство к внешнему поведению? И не является ли эта «точка» просто химерой академической нейрофизиологии?
(Понятие «точка» ни в коем случае не надо понимать буквально, это, скорее, некий лексический образ.)
В своё время сэр Чарльз Скотт Шеррингтон на вопрос о том, что в организме является правящим началом, тем самым «я», что провоцирует и концентрирует вокруг себя (замкнутую в некоей соматической форме) биологическую жизнь, ответил следующим образом: «В многоклеточном организме, в особенности, если речь идёт о высших реакциях, составляющих его поведение, как социальной единицы в экономике природы, именно нервные реакции, par excellence играют интегрирующую роль, объединяют организм в одно целое и образуют из простого набора органов биологическую индивидуальность» (Sherrington С. S. The Integrative Action of the Nervous System, 1906).
Эта классическая формулировка Шеррингтона, разумеется, безупречна, но (как легко заметить) она содержит в себе не ответ на наш вопрос, а ряд новых вопросов, и требует сегодня существенных корректировок и уточнений.
Отметим, что Шеррингтон здесь даже не предполагает наличия некоего «центрэнцефалического начала», некоей «точки» или же отдельной структуры мозга, управляющей как прочими церебральными формациями, так и нервной системой в целом. Он говорит лишь о «нервных реакциях», которые суть производные всей нервной системы.

Илл. 45. Ч. С. Шеррингтон
Вот здесь уже требуются корректировки.
Общая интегрирующая роль нервной системы в любом организме — несомненна, но утрата даже значительной её части, которой может лишиться существо с частью органов (как, к примеру, человек при ампутации конечностей или ряда внутренних органов), не приводит ни к каким существенным изменениям или ослаблениям той функции, что «объединяет организм в одно целое» и создаёт «биологическую индивидуальность ».
Уже одно это обстоятельство позволяет вывести за пределы поисков «интегративного начала» как вегетативную нервную систему, так и часть «завязанной» на неё центральной, существенно сузив круг «обитания» нашей предполагаемой «точки».
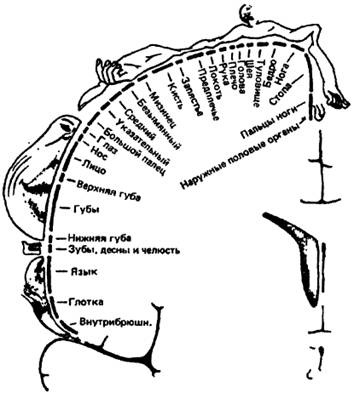
Илл. 46. «Гомункулюс» Пенфилда
К примеру, в области прецентральной извилины установлены центры, «раздражение которых вызывает расслабление или сокращение мышечных оболочек желудка и кишечника» (Лермитт Ж., 1929); В. М. Бехтерев, Миславский, Ч. С. Шеррингтон, Пуесен подтвердили в области парацентральной дольки существование полей, раздражение которых вызывает разнообразные реакции полового аппарата и так далее. Таких примеров можно привести множество, но лучше, говоря о «представительствах» в коре различных вегетативных, тактильных и моторных функций — обратить пристальное внимание на т.н. гомункулюс Пенфилда, хрестоматийную фигурку, достаточно точно демонстрирующую «занятость» коры обеспечением (а возможно, тонкой корректировкой) тех функций, утрата которых драматична для любого существа, но не ведёт к потере «биологической индивидуальности» (илл. 46).
Вероятно, столь же уверенно можно удалить из «круга» наших поисков не только все эволюционно позднейшие церебральные образования, но даже и те древние формации, что имеют строго «служебные» функции обеспечения анализаторов и рецепторов.
Поясняю.
Первую группу («позднейшую») мы «удаляем», согласно пониманию того, что «биологической индивидуальностью», в такой же степени, как и сам сэр Чарльз Скотт, был и любой протокраниат, звероящер или «первоплацентарная» эомайя; т.е. что мозг значительно более ранней «модификации», нежели тот, что предлагает нам анатомия неоцена, бесспорно, уже нёс в себе интегрирующую функцию.
Для лучшего понимания того, каких базовых формаций может лишиться головной мозг, не лишаясь, вероятно, при этом своей важнейшей интегративной функции, приведу опыт полных или частичных децеребраций (удаления мозговых полушарий), проведённых Флюрансом, Мунком, Христиани, Дюссе де Бареном (1902-1919 гг.). Как писал Ж. Лермитт (1929): «К сожалению, это вмешательство было до того серьёзно, что животные погибали через несколько дней». Ознакомление с протоколами децеребраций подтверждают показания Лермитта. В 99% случаев подопытные мучительно умирали, но сам факт их установленных страданий, «мучений, тоски и ужаса», полагаю, наилучшее подтверждение сохранения ими «биологической индивидуальности» до самого момента смерти. Кстати, этот «момент», т. е. непосредственно смерти, прекрасная иллюстрация того смысла, который Ч. Шеррингтон вкладывал в свой термин «биологическая индивидуальность». Смерть — это как раз тот фактор, который ликвидирует, прежде всего, именно её, обезличивая любую биологическую структуру и уравнивая её с любой другой формой неиндивидуализированной органики.
Достойно упоминания, что в 1923 году Макс Ротман (Max Rothmann) произвёл полную децеребрацию собаки, но удалил полушария столь искусно, что оставил интактными все таламические структуры, полосатое тело, мост и нижележащие структуры ствола. (Г. Ротман (Н. Rotmann) и Б. Бруве (В. Brauwer) в этом же году опубликовали полный анатомический протокол эксперимента). В этом случае собака прожила более трёх лет, но в состоянии «поразительного уменьшения деятельности всех органов чувств».