(Stark P, Boyd E. Effects of Cholinergic Drugs on Hypothalamic Self-Stimulation Response Rates of Dogs, 1963; Goodman I., Brown J. Stimulation of Positively and Negatively Reinforcing Sites in the Avian Brain, 1966; Lilly J., Miller A. Operant Conditioning of the Bottlenose Dolphin with Electrical Stimulation of the Brain, 1962; Persson N. Self-Stimulation in the goat, 1962.)
Человек лишь шесть раз стал объектом таких экспериментов (К. Сэм-Якобсен и А. Торкильдсен; М. Бишоп — С. Елдер), и во всех этих случаях эффект, открытый Д. Олдсом и П. Милнером был (в общих чертах) успешно повторен.
(Полагаю, исследования на homo не могут быть признаны безупречными или даже полноценными. Хотя электрораздражение проводилось с согласия пациентов и лишь сопровождало нейрохирургические операции, т.н. этические нормы существенно ограничивали возможности исследователей.)
(Sem-Jacobsen С, Torkildsen A. Depth Recording and Electrical Stimulation in the Human Brain, 1960; Bishop M., Elder S., Heath R. Intracranial Self-Stimulation in Man, 1963.)
Несмотря на обилие лабораторного материала и титулованность кафедр, где повторялось, усложнялось и совершенствовалось это открытие, мы всё же вернёмся в скромную лабораторию Хэбба, к первому эксперименту Д. Олдса и П. Милнера.
Как вы, вероятно, помните, смысл опыта заключался в том, что подопытной белой крысе с вживленными в мозг электродиками была предоставлена полная возможность раздражать определенные зоны своего мозга через нажатие педальки в несколько видоизменённом «ящике Скиннера» (илл. 58).
«Раздражающие электроды, хронически вживлённые в подкорковые структуры (мозга крысы — Прим. автора), были соединены посредством тонкого гибкого провода с источником тока» (Окс С. Основы нейрофизиологии, 1969).

Илл. 58. Опыт Дж. Олдса — П. Милнера
Впрочем, самое лучшее описание своего опыта даёт сам Олдс:
«Когда крыса наступает на педаль (мы использовали самые различные виды педалей), она производит раздражение собственного мозга. Раздражение прекращается через полсекунды после включения, даже если крыса продолжает нажимать на педаль; поэтому для нанесения повторного раздражения крыса должна отпустить педаль и снова нажать на неё. Если во время самораздражения электрическая цепь прерывается экспериментатором, так что надавливание на педаль перестает вызывать раздражение, животное несколько раз с яростью нажимает на педаль и только после этого отворачивается от педали и начинает чиститься или засыпает... В качестве поощрения при надавливании на педаль крыса всегда получает лишь раздражение мозга...» (Olds J. Differentiation of Reward Systems in the Brain by Self-stimulation Technics, 1960).
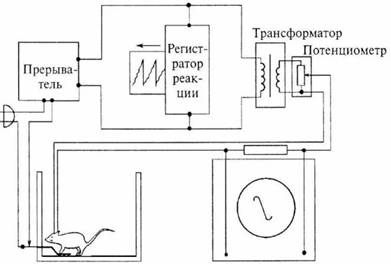
Илл. 59. Схема опыта Олдса.
Нажимая на рычаг, крыса на короткое время включает электрический
ток, который вызывает раздражение её мозга (самораздражение).
Реакции на нажатие регистрируются специальным прибором; сила
раздражения измеряется с помощью осциллоскопа
Примечательно, что олдсовская белая крыса, ради получения тех ощущений, которые давали разряды синусоидального тока 5-100 μa с частотой 60 Гц, пренебрегала любыми другими радостями жизни, т.е. сном и едой. Количество нажатий и, соответственно, полученных мозгом крысы разрядов, достигало 5000 в 1 час. (Иногда — до 8000) (илл. 59).
«В опытах на крысах Олдс установил, что животные могут нажимать на рычаг безостановочно, до полного изнеможения. Животные предпочитают самораздражение приему пищи, спариванию; они готовы испытать боль с тем, чтобы добраться до рычага и возобновить самораздражение» (Окc С. Основы нейрофизиологии, 1969).
К. Сем-Джекобсен и А. Торкильдсен во время нейрохирургических операций на ненаркотизированных больных людях решились (отчасти) повторить эксперимент Олдса, током той же, «крысиной», частоты (и электродом того же калибра, т.е. 0,25мм) раздражая различные участки мозга своих пациентов. Повторяю, надо понимать, что в этих случаях опыты не могут считаться полноценными, так как исследователи были сильно ограничены во времени, в количестве раздражаемых точек, в глубине установки электродов. Тем не менее «получив возможность контролировать частоту раздражения, испытуемый прибегал к самораздражению... Больной отмечал появление чувства радости и удовлетворения, а временами — приятное щекотание в мышцах области таза» (Sem-Jacobsen С., Torkildsen A. Depth Recording and Electrical Stimulation in the Human Brain, 1960). На все вопросы о возможных различиях реакций, связанных с различием мозга homo и rattus norvegicus, ещё в тридцатые годы XX века ответил классик нейрофизиологии, радикал-академист Карл Спенсер Лешли (1890-1958), который в своих экспериментах 1909-1929 гг. имел возможность доскональнее прочих разобраться в структурах и особенностях мозга лабораторных крыс: I. «Я начинаю сомневаться в том, что эволюция млекопитающих привела к какому-либо изменению основной организации или механики деятельности головного мозга. Наблюдающиеся в этом отношении большие различия являются скорее различием в объёме, чем в организации» (The Relation Between Cerebral Mass, Learning and Retention, 1925); II. «Помимо функции пространственной ориентации, имеется мало данных, которые бы свидетельствовали о более тонкой корковой дифференциации у человека в сравнении с крысой» (Мозг и интеллект, 1933).
Уточним топографию вживления электродиков в опытах Д. Олдса и его последователей.
Разумеется, в первую очередь воздействиям тока подвергались древнейшие структуры обонятельного и среднего мозга: гипоталамус, гипоталамические структуры, амигдала, гиппокамп, энторинальная зона, свод, перегородка, а также различные участки, где находятся средоточия ядер ретикулярной формации.
Следует отметить, что наивысшей интенсивности (7000-8000 раз в час) самораздражение достигало при установке электродов в область nucleus interpeduncularis, которое относится к бесспорным (даже по Бакстеру и Ольшевскому) ретикулярным ядрам, а судя по его характерному расположению, возможно, к ядрам наидревнейшим, с которых и началась когда-то история головного мозга.
Интерпедункулярное ядро ретикулярной формации (nucleus interpeduncularis) мы можем без труда обнаружить у миксины, миноги, акулы и крокодила. У миксины (согласно Л. Эдингеру, 1911) оно имеет ещё несколько размытую форму, но уже у миноги получает первую отчётливость очертаний; у акулы являет собой почти полноценное ядро и хорошо наблюдается аккурат под ядрами глазодвигательного нерва, чуть кзади от fasciculus longitudinalis centralis (по Капперсу), а вот у крокодила обретает уже все свое окончательное великолепие и становится почти неотличимо от nucleus interpeduncularis медведя, человека или свиньи. (У млекопитающих оно в полном соответствии со своим названием располагается над межножковой ямкой.)
Полагаю, необходимо упомянуть и то, что электровоздействие на ряд «точек» в архео- и палеоструктурах вызывало не некую «общую», а лишь локальные специфические реакции: семяизвержение, сильную жажду, голод, сон (Milner Р. Physiological Psychology, 1970). Но таких точек оказалось всё же значительно меньше, чем зон «общего» возбуждения мозга, и наличие точек специфического возбуждения никак не меняло глобальной картины.
Столь же существенен и тот факт, что частота самораздражения уменьшалась по мере удаления электродиков от ядер ретикулярной формации, а в некоторых областях прекращалась вовсе (район свода и зубчатой извилины), что вполне могло быть указанием либо на дискомфортность, либо на прямую болезненность воздействия тока.
Но при всех экспериментах твердо фиксировалось значительное преобладание зон, воздействие на которые провоцировало бесконечные повторения: «Только при раздражении некоторых участков этой системы можно наблюдать стремление животного избегать электрического раздражения. Следовательно, у крыс область раздражения, которой животное избегает, чрезвычайно мала по сравнению с обширной областью, к электрическому раздражению которой животное стремится» (Olds J. Differentiation of Reward Systems in the Brain by Self-stimulation Technics, 1960).
Сумма этих фактов, неоднократно подтвержденных и зафиксированных, позволила Д. Олдсу сделать чрезвычайно важный вывод: «Таким образом, нельзя рассматривать мозг как систему, стремящуюся уменьшить собственное возбуждение, так как значительная его часть проявляет тенденцию к увеличению своего возбуждения» (Olds J. Differentiation of Reward Systems in the Brain by Self-stimulation Technics, 1960).
Хотя нет никаких оснований сомневаться в постулате У. Г. Пенфилда: «Рефлекторная деятельность, несомненно, совершенно одинакова у человека и высших животных» (Пенфилд У. Г. Эпилепсия и функциональная анатомия головного мозга человека, 1958), да и добросовестность «крысиных» экспериментов является очевидной, но олдсовский опыт нельзя безоговорочно переносить на исследуемую нами гипотезу о мышлении как о некоем глобальном или «универсальном» способе самораздражения мозга.
Результаты опытов Олдса лишь дают известное право говорить о «жадности» мозга к различным раздражающим факторам и о том, что продуктом этих раздражений могут быть весьма любопытные процессы.
Мышление, вероятно, лишь один из таких процессов, но мы сильно переоцениваем его «глобальность» и его значимость для мозга. Прежде всего потому, что оно явно (и значительно) уступает по своей раздражительной силе как тем воздействиям, которые были опробованы в лаборатории Хэбба, так и многим другим.
Оно, несомненно, обладает определённой раздражительной силой, но эта сила не так велика, чтобы сделать процесс мышления самым «желанным» и глобальным средством возбуждения церебральных структур.
Что же может служить самым простым индикатором экстремальности или «штатности» физиологического состояния этих структур?
Прежде всего — ликвор, т.е. питающая и омывающая мозг цереброспинальная жидкость, являющаяся и «кормильцем» мозга, и его основной «средой».
«Ликвор является зеркалом, отражающим процессы, происходящие в тканях мозга» (Фридман А. П. Основы ликворологии: учение о жидкости мозга, 1971).
Однако, судя по неизменности давления, химизма, объёма, электропроводимости цистернального и вентрикулярного ликвора, т.е. по относительной физиологической «безмятежности» мозга, мышление в «масштабах» мозга не является не только экстремальным, но даже и просто «заметным» явлением.
Известно, что любые колебания физиологического состояния мозга в первую очередь отражаются на химизме, давлении, электропроводности и количестве ликвора. Гнойный и эпидемический менингиты, полиомиелит, миелит, табес, любые сосудистые заболевания мозга — резко меняют данные нормы электропроводимости, т.е. с 1,05 × 102 — 2,00 × 102 (по Фридману) до самых различных значений. Эпилептоидные состояния, алкоголь, люэс, малярия резко меняют как электропроводимость, так и химизм ликвора (Klemperer et Weismann, 1940). Не столь резко, но всё же существенно состав ликвора (щелочной) изменяют даже такие факторы, как сон или бессонница (Lejandr et Pierr, 1921); а изменение температуры тела или простейший эфирный наркоз кардинально искажают в нём т.н. картину сахара.
Психические заболевания влияют на содержание в ликворе дофамина, серотонина, ГАМК, кальция (Цветанова, 1986). Более того, на ликворное давление оказывает сильное влияние даже факт общей вертикализации тела (стояния) или полувертикализации (сидения). Но никаких данных о метаморфозах ликвора соотносительно с процессом даже самого напряженного мышления не существует. Картинки КТ, МРТ и даже ПЭТ хороши для формальной диагностики гематом, гидроцефалий, вентрикулитов и так далее, но никак не для оценки «процессов мышления». (Полагаю, для обсуждения «открытий-однодневок» или откровенных спекуляций здесь не самое подходящее место.)
Ещё одним косвенным доказательством того, что мышление относится к безусловно раздражительным процессам, но не к экстремальным, а к штатным и, вероятно, мало «обременительным» для мозга, может служить сопоставление электроэнцефалограмм коры решающего математические задачи А. Эйнштейна и кошки, ретикулярная формация которой подверглась одиночному электрораздражению (в области таламуса). И в том и в другом случае мы сперва видим примерно схожий, очень отчётливый α-ритм, прерываемый неким небольшим скачком, характерным для «реакции пробуждения».
С кошкой всё понятно, в момент «скачка» она получила разряд в область таламуса, который немедленно потребовал «ответа» от анализаторов коры.
С А. Эйнштейном все несколько иначе, но всё же, в принципе, сопоставимо и по причинности, и по электроэнцефалограмме. Г. Джаспер, проводивший его электроэнцефалографирование, пишет следующее: «Например, когда Эйнштейн производил довольно запутанные математические операции, которые он, однако, выполнял совершенно автоматически, у него регистрировался отчётливый непрерывный α-ритм. Внезапно α-ритм у него исчез, а сам Эйнштейн казался обеспокоенным. Когда его спросили, случилось ли что-нибудь неприятное, он ответил, что нашёл ошибку в расчётах, которые сделал накануне, и попросил тут же позвонить по телефону в Принстон» (Джаспер Г. Электрофизиология и экспериментальная эпилепсия // Пенфилд У., Джаспер Г. Эпилепсия и функциональная анатомия головного мозга человека, 1958).
Разумеется, речь не идёт об идентичности электроэнцефалограмм или реакций мозга, а лишь о том, что как в том, так и в другом случае все колебания (сравнительно даже с легчайшими ишемическими или эпилептоидными состояниями или реакциями на введение метразола, эзерина, ацетилхолина, стрихнина и так далее) находились в пределах усредненной относительной нормы.
Исследования, специально посвященные электроэнцефалографическому «рисунку» мышления, почти всегда (с незначительными отклонениями) регистрируют обычные колебания альфа-ритма, лёгкое усиление бета-активности, чуть (всплесково) меняясь при добавлении в процесс мышления различных эмоциональных компонентов (Ланге Дж., Сторм Ван Левен В., Верре П. Корреляция между психологическими и электроэнцефалографическими явлениями // Электроэнцефалографическое исследование высшей нервной деятельности, 1962).
Вернёмся к Дж. Олдсу и его экспериментам.
Кстати, при всей уникальности опытов как самого Олдса, так и его последователей история, начавшаяся в лаборатории Хэбба, не получила ни продолжения, ни развития, ни завершения.
Она оказалась лишь подобием «дудочки крысолова». Десятки физиологов во всём мире завороженно повторяли опыт с электрическим самораздражением, но возникающие на его основании гипотезы — умирали в момент их рождения.
Так произошло с «оперантной» теорией самого Д. Олдса (1960), с версией Е. Грастиана (1968) о «возвратном торможении пирамидных клеток гиппокампа», с предположением С. Гликмана и Б. Шиффа (1967) о наличии «биологической системы подкрепления», с концепцией Д. Дэйча (1960) о «мотивационной системе» и так далее (всего было, если не ошибаюсь, не менее 12 гипотез). Тупиковость любой из этих версий была настолько очевидна, что олдсовский эксперимент, как не получивший никакого вразумительного объяснения, в результате был просто забыт нейрофизиологией. Мы не найдем даже упоминания о нём в любом из авторитетных сводных трудов недавнего времени. Ни у Г. Шеперда (1983), ни у Д. Николлса, А. Мартина, Б. Валласа, П. Фукса (1975) — о крысиной истории нет уже ни единого слова, хотя никто и никогда не ставил под сомнение научную добросовестность экспериментов с самораздражением и их впечатляющий наглядный результат.
Вероятно, «тупиковость» гипотез была изначально предопределена некорректностью формулировок, употребляемых как самим Д. Олдсом, так и всеми его последователями.
Обратим внимание на следующий факт.
Даже в протоколах опытов широко и концептуально употребляются такие понятия, как «удовольствие», «центр удовольствий», «поощрение», «подкрепление» и другие термины, не только не имеющие никакого нейрофизиологического смысла, но и вносящие в суть происходящего существенные искажения.
Как мы помним, Иван Петрович Павлов категорически не рекомендовал при трактовке экспериментов переходить на «чужой» язык, «чужие» понятия, каковыми являются любые термины из арсенала психологии: «До сих пор этот отдел в значительной своей части пользовался чужими понятиями — психологическими понятиями. Теперь получается возможность вполне освободиться от этой крайне вредной зависимости. <...> Итак, физиолог в данном пункте оставил твёрдую естественнонаучную позицию. И что он приобрёл вместо неё? Он взял понятия из того отдела человеческого умственного интереса, который, несмотря на свою наибольшую давность, по заявлению самих его деятелей, не получил ещё до сих пор права называться наукой» (Павлов И.П. ПСС, 1951. Т. Ill, кн. 1).
К слову, Ч. С. Шеррингтон тоже обращал внимание на недопустимость и бессмысленность, как он выражался, «поддельной» или «псевдоэффективной» терминологии (Woodworth R. S., Sherrington C. S. A Pseudoaffective Reflex and its Spinal Path, 1904).
Приведем простой пример того, как неточные, чужие термины способны исказить и даже обессмыслить восприятие физиологического процесса. Это несколько отвлечет от темы, но я постараюсь быть предельно краток.
Ознакомившись с описаниями опытов, сделанными самим Дж. Олдсом, мы обнаружим следующие, полагаю, абсурдные формулировки: «крыса раздражает свой мозг», «животное совершает раздражение своего мозга», «когда крыса наступает на педаль, она производит раздражение собственного мозга» (Olds J. Differentiation of Reward Systems in the Brain by Self-stimulation Technics, 1960).
Рассмотрим протокол другого подобного эксперимента и вновь обнаружим: «Обезьяна научилась замыкать контакт триггера с частотой этих сокращений и производила до 18 раздражений своего мозга в 1 сек. Обезьяна действовала таким образом в течение 20 часов до полного изнеможения, производя около 200 000 раздражений» (Lilly J. Learning Motivated by Subcortical Stimulation, I960).
Однако обратимся и к самому академичному из описателей, к Питеру Милнеру, но и у него «раздражение мозга может доставить животному удовольствие», «кошки, обученные нажимать на рычаг, чтобы получить раздражение мозга» (Milner Р. М. Physiological Psychology, 1970).
В отношении прочих существ (коз, дельфинов, собак, голубей, людей), на которых проводились опыты с самораздражением, исследователями употреблялась сходственная фразеология.
Легко заметить, что в соответствии с этими формулировками и у П. Милнера, и у Дж. Лилли, и у Дж. Олдса в их опыты вовлечены какие-то необычайные, волшебные существа «крыса», «обезьяна», «кошка», т.е. некие вненаучные «образы», распоряжающиеся собственным мозгом для получения «удовольствия».
В данном случае — это не мелочная придирка к «вопросам стиля» или удобствам номинаций; это констатация типичного «переворота» понятий и той некорректности, что, вероятно, и включает известный principium ludi domini (принцип игры господина?), при котором факты обрушивают друг друга, заводя в тупик любую гипотезу.
Вероятно, следует уточнить, что «крыса», «кошка», «дельфин», «коза», «обезьяна» или «человек» (у Сэм-Якобсена и Торкильдсена) к самораздражению никакого отношения не имели. Самораздражение производили сходственные у всех этих животных структуры мозга, а уж какое именно «тело» использовалось ими для обеспечения этого воздействия, крысиное, человеческое или кошачье, принципиальным не является.
Тут следует отметить, что понятие «удовольствие» при всей его расплывчатости сильно разнится для столь морфологически несхожих существ, как крыса, кошка, обезьяна, голубь, коза, homo или дельфин.
В этом контексте становится понятна малая уместность терминов «поощрение», «подкрепление», «удовольствие» и настоятельная необходимость очистки результатов опыта Дж. Олдса от «словесной шелухи».
Кроме того, схема самого Дж. Олдса не оставляет сомнений в том, что раздражению подвергались те области, что при штатных воздействиях среды генерируют отнюдь не «блаженство», а различные агрессии, активность ассоциативной «базы» гиппокампа, кортикопетальные и кортикофугальные связи, моторику и так далее (илл. 60).
Стоит отметить:
I. Что физиологические, эмоциональные, адаптивные состояния крыс (или иных подопытных) при электрораздражениях не сходствовали с последствиями применения к ним фармакологических препаратов наркотизирующего, депрессирующего или возбуждающего типа.
II. Сочетание электрораздражения и фармакологии каждый раз имело разные последствия: хлорпромазин, имипрамин, эзерин, резерпин подавляли ритм нажатий на педальку; атропин, мепробамат и пенобарбитал не оказывали заметного влияния; амфетамин способствовал незначительному учащению ритма (особенно у коз в опытах Перссона (1962)).
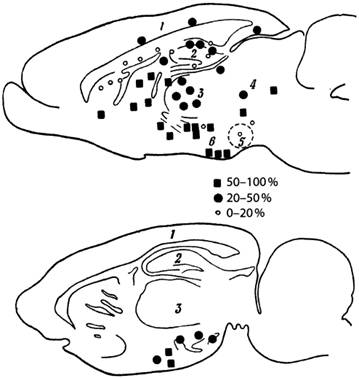
Илл. 60. Парасагиттальные сечения мозга крысы,
показывающие размещение участков самораздражения
1 — кора; 2 — гиппокамп; 3 — таламус; 4 — покрышка; 5 — мамиллярные тела;
6 — гипоталамус
Указанные процентные показатели вычислены на основании учёта периодов длит. 30с и более, в течение которых реакции самораздражения отсутствуют (по Милнеру)
В этом строгом контексте мы остаёмся наедине лишь с фактом стремления стволовых структур к гораздо более мощным возбудителям, чем те, что может предложить среда в её обыденном варианте; мы, вероятно, видим те подлинные рефлекторные потенциалы древних отделов мозга, что сформировались в течение 500 миллионов лет непрерывной эволюционной драмы.
Грубо говоря, ни на какие иные выводы эксперименты Дж. Олдса не дают никакого права, а сведение их результатов к различным «поощрениям» и «удовольствиям» — это лишь попытка подменить литературными терминами не очень понятные рефлекторные процессы, в нюансах которых ещё предстоит разбираться будущим поколениям нейрофизиологов. Впрочем, и то немногое, что представляется нам несомненным на данное время, тоже является существенным для нашего исследования, т.е. огромный рефлекторный потенциал и «жадность» мозга к сильным, а по всей вероятности — и к любым раздражителям.
Здесь стоит мимоходом, но внятно отметить, что ещё в конце XIX столетия, не имея никаких экспериментальных данных, а основываясь лишь на понимании логики мозга и его эволюционной истории, И. М. Сеченов высказал предположение, «что нервная система обладает потребностью в действии, т. е. она должна иметь определенный минимум возбуждения». Это положение чуть позже, но им же было развито в известном утверждении, где И. М. Сеченов почти впрямую аналогизирует «мышление» и «ощущение»: «Следовательно, если смотреть на ассоциацию только в отношении ряда центральных деятельностей, то она есть непрерывное ощущение» (Сеченов И. М. Избр. произв.: в 2 т. Т.1. Физиология и психология, 1952).
Заслуга Дж. Олдса в том, что он обнаружил «пиковую точку», возможно, даже некий апогей рефлекторной активности мозга, попутно доказав, что стремление к биологическому покою, к «экономности» рефлекторных ответов не является потребностью головного мозга, а, что, напротив, у мозга существует отчётливая тенденция к эскалации своей активности.
Сегодня можно лишь предполагать, в чём причина этого физиологически парадоксального свойства мозга? Кто является истинным «автором» этой эскалации: свойства ли самих нейронов, особенности их связей или активирующие мозг древние стволовые формации?
Вероятно, если желание решить этот вопрос всё же возникнет, то противоборствовать будут именно «ретикулярная» и «синаптическая» гипотезы.
К слову, следует отметить, что того яростного, подлинного интереса к работе механизмов мозга, которым был отмечен период с последней четверти XIX столетия до последней трети XX, уже не наблюдается, а возрождение его в ближайшем будущем не просматривается. Тот минимальный интерес к данному вопросу, что ещё существует, легко удовлетворяется формальными картинками МРТ или интересными, но очень уязвимыми теориями о всесилии «синаптических связей». (Без почтительного упоминания о которых не обходится сегодня ни одно исследование о мозге, как, впрочем, в 80-е годы не обходилось без славословий забытой ныне «нейрокибернетике».)
Версия генерации общей и частной активности мозга или того, что лидер данного направления Э. Кэндел называет «памятью» именно в синаптических связях, несомненно, крайне любопытна (Kandel Е. In Search of Memory, 2006).
Кстати, ещё задолго до своего «звёздного», «нобелевского часа» она была уже известна, дискутирована и с позиций классической нейрофизиологии подвергнута сомнению: «Было бы наивно думать, что следы памяти могут храниться “in toto” в закодированных структурах молекул, в отдельных нервных клетках или синаптических структурах нейрональных мембран. Биологические процессы роста и обучения зависят от взаимодействия между отдельными элементами в организованных популяциях клеток, находящихся под влиянием внутренних и внешних стимулов и окружающей среды» (Джаспер Г. Механизмы отбора и хранения приобретённых форм стимул-реакции // Современные проблемы электрофизиологии центральной нервной системы, 1967).
Г. Джаспер здесь вежливо, но тщательно подчеркнул традиционную для «радикального академиста» уверенность в ошибочности переноса поиска понимания механики мозга — в область клеточных и межклеточных микропроцессов.
Э. Флори (1972) сформулировал свое видение «синапсизма» чуть мягче, но, по сути, ещё точнее обозначил, что искать возможность объяснения полноценной памяти, интеллекта и мышления homo на уровне синаптических связей как минимум не слишком продуктивно: «Действительно, основные свойства синапсов одинаковы во всём царстве животных. Мы находим те же принципы строения в синапсах кишечнополостных и в синапсах млекопитающих: нервные окончания всегда содержат митохондрии и синаптические пузырьки, синаптическая щель имеет те же размеры, даже медиаторы и связанные с ними энзимы, возможно, являются одинаковыми» (Эволюционные аспекты синаптической передачи // Эволюционная биохимия и физиология, 1972. Т. 8).
Впрочем, дело даже не в том, что положения авторов «торжествующего синапсизма» конфликтуют с классической нейрофизиологией.
В конце концов всё в той или иной степени релятивистично, догмы условны, а свобода сомнений — безгранична и обязательна.
Но при всей важности этих исследований «синапсизм» подобен попытке выяснения принципов работы сейфового механизма через анализ молекулярного состава стали, из которой сделаны ригели, сувальды и пружинки замка.
Ведь даже сверхточно выяснив расположение всех атомов в кристаллической решетке этой стали, мы всё равно ничего не узнаем о том, как работает механизм сейфа, ибо его работа производится разделенными, взаимодействующими и геометрически строго оформленными структурами, порождёнными средой и ей подчинёнными.
Эта аналогия грубовата (на грани некорректности), но подразумевает лишь то, что полиструктурность мозга есть безусловный факт, что в известной степени специализация его анатомических структур понятна, а их взаимодействие очевидно.
К слову, надеюсь, что в качестве примера не надо приводить миллионы нейрохирургических операций, успех которых основывается прежде всего на точных представлениях о локализации различных функций в различных мозговых структурах. Достоверность этих представлений настолько высока, что позволяет хирургу при удалении опухолей, кист и гематом «не задеть» те точки, мельчайшее повреждение которых приводит к афазиям, коме или иным тяжким для пациента последствиям.
Если бы синаптическая или иные подобные версии (из тех, что сводят работу мозга к микропроцессам в везикулах, к пропуску ионов через мембраны, к активациям протеинкиназы A и так далее) годились бы для объяснения церебральной работы, то даже полушарность для такого мозга была бы излишней и необъяснимой эклектикой; этот орган вполне мог бы быть анатомически бесструктурным «клеточным монолитом», организованным значительно проще, чем даже печень. Наличие в нём структур, формаций, локализации функций, анатомически наблюдаемых аксонных пучков, комиссур и лучистостей — всё было бы ненужным и бессмысленным.
Сегодняшнее противостояние догм классической нейрофизиологии и «синапсизма» отчасти напоминает ситуацию двухтысячелетней давности: вспомним спор Галена Пергамского с «аристотелевцами», утверждавшими, что мозг монофункционален и служит лишь для «охлаждения сердца». Гален тогда задал весьма логичный вопрос: «Если бы мозг оказался созданным только для охлаждения, то зачем тогда существуют различные его части, такие как сосудистые тела, сетевидные сплетения, шишковидная железа, воронка, гипофиз, псалтыревидное тело (Судя по всему — комиссура гиппокампа, которая в совсем старых анатомических фолиантах именуется как «лира Давида» — Прим, автора), червеобразный отросток? Для охлаждения было бы достаточным, если бы головной мозг был сделан наподобие бездеятельной и бесформенной губки, и тогда не было бы нужды в столь искусном строении, какое мы находим в мозгу» (Цитата Галена приводится по «Трактату о функциях нервной системы» Г. Прохазки (1784)).