Для того чтобы определить экспрессию генов паразита, как он относится к развитию Toxoplasma в том-опосредованном хозяине, SAGE библиотека (Velculescu и др., 1995) были построена из Вег деформации популяций-ций спонтанно возникающих из спорозоитов Infec-ных человеческих фибробластов (Jerome и др., 1998), и из лабораторных штаммов адаптированный, представляющих основные генотипических варианты Toxoplasma (тип I-RH, тип II-Me49B7 и типа III-VEGmsj). библиотеки SAGE были построены с использованием источников мРНКа, полученные из ключевых переходов в разви-opmental программы из спорозоитов в брадизоитном, как описано в более ранних исследованиях (Иероним и др., 1998). В общей сложности 290 000 тегов были получены с помощью этой Proj-ЭСТА и индивидуумы могут получить доступ к полному набору данных на TgSAGEDB (https://vmbmod10.msu.montana.edu /vmb/white-lab/newsage.htm) или SAGE просматривать данные в toxoDB-GBrowse (HTTP: // toxodb.
16.2.3.1 SAGE глобальная статистика и хромосома пейзаж
В ~ 300 000 метки, генерируемая проект SAGE представляет 8880 уникальных последовательностей 14, что мономерных

422 GENE РЕГУЛИРОВАНИЕ
матч токсоплазмы последовательность генома с ограниченной избыточности, предполагая, что почти 9000 уникальных транскриптов мРНК выражены в токсоплазмы в течение промежуточного жизненного цикла. По оценкам, 6300 генов (на основе CAP3 сборки 2-т.п.н. геномных последовательностей 5'-upstream каждого тэга) представлены эти метки, причем избыточность объясняется дифференциальной полиА+выбор и потенциальные варианты сплайсинга. Кроме того, проект SAGE определил мРНК, содержащий единичные нуклеотидные полиморфизмы, а также некодирующие РНК предст-senting потенциальных стенограмм антисмысловых. Среднее содержание тегов SAGE хромосомы было замечание, умело согласуется с меткой SAGE для каждого 6000 п.о. во всех хромосомах. Таким образом, примерно половины генома токсоплазмы занимают ген-тран-scription единиц с относительным расстоянием генов ~ 2000 п.о. (с использованием средней ~ 4000 п.о. за ген, полученный из целого генома аннотации, Макку и Roos, неопубликованные данные). Ограничения расстояния являются общей чертой промоторов в одноклеточных эукариот с компактными геномов (Stamatoyannopoulos, 2004), а также несколько Toxoplasma промоутеров изучены в соответствии с этой структурной организации - например,
Почти 49 процентов SAGE теги из Вег Devel-opmental этапов показывают> 2,5-кратное изменение частоты в по меньшей мере одной библиотеке (66 процентов> 2 раза), что свидетельствует о том, что регуляция транскрипции обширна в этом паразита. Классы мРНК обилие, как это определено тремя частотами тегов SAGE (Velculescu и др., 1995) - высокие> 100, умеренные 11-99, и низкий уровень - 2-10 разбиты на 70 процентов низкое содержание тегов, 25 процентов умеренных тегов и 5 процентов высокого содержания метка. По сравнению с дрожжами (Velculescu и др., 1997), с высоким и средним уровнем численности бассейны меньше в этом паразита (половина размера дрожжей), что указывает на менее конститутивной экспрессии гена в Toxoplasma (рис 16.1). В отличие от высокой экспрессии мета-Болич или структурных генов в клетках животных, почти
треть Toxoplasma-обильное мРНК группы являются генами Apicomplexa специфичного, которые имеют простые геномные структуры, содержащие мало, если какие-либо интроны (основано на частоте, один из пяти SAGE теги Corre-ответствует жестко регулируемые Apicomplexa-специфические транскрипты из плотных гранул, НКГА, МИК, УОП; Рисунок 16.1). Таким образом, многие транскрипты, кодирующие белки базальный метаболический Мачин-Эри и субклеточных структур по всей видимости, Тран-скрайбированной в токсоплазмы только при необходимости в процессе роста и развития паразита. Это consis-палатка с концепцией «точно вовремя», выдвинутой из исследований Plasmodium (Ллайнас и DeRisi, 2004). В целом, гены развития специфического (спорозоиты, тахизоит, брадизоитные), гены, кодирующие белки из биохимических путей, а также гены, представляющих мРНКи классов обилия рассредоточены между всеми хромосомами Toxoplasma.
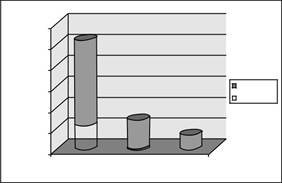
| 66% | |||||
| подсчитывать | |||||
| Всего теги | |||||
| Тег | Api Specfic | ||||
| 22% | |||||
| 11% | |||||
| > = 100 | 11--99 | <= 10 | |||
РИСУНОК 16.1 Высокого класса обилие мРНК в Toxoplasma обогащена Apicomplexa-специфических генов. Нормированные теги SAGE были разобраны на три класса в зависимости от частоты. Теги, найденные в> 100 были рассмотрены высоко в изобилии, в то время как метки с частотой от 99 до 11 или <10 были умеренно или низко в изобилии, соответственно. На основе этих классов частот ~ 270 уникальных транскриптов охватывает 66 процентов всех тегов в общем проекте SAGE. Один из пяти тегов SAGE кодирует Apicomplexa-специфического гена, участвующего в передаче паразита и инвазии - например, белки в micronemes, rhoptries, плотных гранул и поверхностных антигенов (56 582 были из Апи-тегов с> 100 Aver-возрастной частоте, 1999 были из 99-11 частот и 426 метки были из <10 частоты). Количество уникальных умеренных тегов составило 1541, и 6286 уникальных тегов имели низкую частоту изобилия.
| Транскриптома токсоплазменных |
возникают и в тех случаях, паттерны экспрессии мРНК не по всей видимости, сильно зависит от физической близости. Так, например, гены ENCOD-ные енолазы 1 и 2 меньше, чем 1500 п.о. друг от друга на хромосому VIII, но выражены исключительно в брадизоитных или тахизоят стадии, соответственно (Lyons и др., 2002). Эти наблюдения указывают на то, что локальные изменения структуры хроматина или набор РНК-полимеразы промоутеров оказывает незначительное влияние на соседние гены. В целом, формирующийся вид организации генов и глобальной экспрессии мРНК, представленной в рамках проекта SAGE указывает, что транскрипционные механизмы играют ключевую роль в регулировании программы развития как токсоплазмы; и общие черты паразита транскрипции мРНК (например,
16.2.3.2 SAGE анализ первичных паразитов VEG штамма из спорозоитов в тахизоит к брадизоитному в сравнении с лабораторными штаммами
Глобальные сравнения SAGE теги из Вег первичных библиотек (спорозоитами, 4-й день, 6, 7, 15 после спорозоитов инфекции и рН-сдвига источники мРНК используются для построения первичных библиотек SAGE, Иероним и др., 1998) показывают уникальные изменения уровня мРНК определение каждого из этапов развития (корреляции Пирсона г çíàê ðàâíî 0.0229-0.32), за исключением спорозоитов и паразитов, выходящих из спорозоитами инфекций на 4-й день (г çíàê ðàâíî0,728). SAGE теги однозначно связаны с каждым развитием, умственные библиотеками охватывают 23 процентов от общего количества тегов секвенирован, и варьировались от 1.5-5.3 процента тегов в каждой библиотеке. В мировом масштабе, наши результаты параллельно аналогичные исследования в Plasmodium, что инди-Cate механизмы координации с развитием экспрессии мРНК в этих паразитов в значительной степени конкретной стадии. Кроме того, обнаружение больших групп стадии специфических генов в Toxoplasma добавляет поддержку концепции, что с развитием генной экспрессии в этом паразита, вероятно, следует за Шегаг-chical порядка (Синг и др., 2002). Таким образом, наши исследования SAGE (и более ранние эксперименты микрочипов; Singh
и другие., 2002) свидетельствуют о том, что мРНК Експрес-Sion в Toxoplasma падает в различных совместных регулируемых классов и групп мРНК, которые, вероятно, регулируемые разработки конкретных факторов транс-действующих, управление фенотипические переходы, связанные с промежуточным жизненного цикла токсоплазмы.
Анализ экспрессии гена через развитие паразита показал ожидаемые паттерны экспрессии мРНКа, а также удивительных профилей мРНКа, которые могут обеспечить новое биологическое понимание генетической программы, выраженной Toxoplasma, и помогают объяснить вариации в способности развития, которые происходят между штаммами. Мы сообщали, что паразиты, выходящие из первичных спорозоитов прививок (VEG деформации) подвергаются ~ 20 быстрых делений прежде, чем синхронно замедление скорости их ростов и инициирование брадизоитной экспрессии гена приблизительно 1 недели после спорозоитов инфекции (Иероним и др., 1998). Таким образом, это не было неожиданным, что мы наблюдали в понижающей регуляции частоты тегов SAGE, соответствующих генов роста, связанных (например, TgPCNA1, DHFR-TS, и аденозин транспортером) и некоторых связанных с энергией генов (например, фруктозо-1,6-фосфат, gyceraldehyde-3-фосфат, 3-phosphglycerate) в библиотеке День-7 (и все последующие библиотеки сдвига после роста) конгруэнтны с существенно медленнее, время удвоения этих паразитов. Далее согласен-ния с более ранними измерениями BAG1 в этих популяциях (2 процента BAG1 на 7-й день после СПОРО-zoite;. Иероним и др, 1998) было отражено в отсутствии известных генов брадизоитный в этой библиотеке. Паразиты от Day-15 после инфекции спорозоитов представляют собой смесь тахизоит и брадизоитный форм (50 процентов BAG1;. Иероним и др, 1998), и, таким образом SAGE метки, соответствующие брадизоитный маркеры SAG4.2 и ENO1 были очевидны в этой библиотеке наряду с выражением тахизоит-специфических генов, таких как SAG1. Воздействие Векли первичные паразит до рНа стресса сдвигает бассейн мРНКа дополнительно к стадиям брадизоитных (Радка и др., 2003), так как все из опубликованных брадизоитных маркеров были обнаружены (Lyons и др., 2002), и имели более высокие частоты, в рНе смещенной библиотеки SAGE, чем наблюдались в библиотеке дня-15. Не менее важно, и предсказал, было практически полное подавление генов тахизоят конкретные
424 GENE РЕГУЛИРОВАНИЕ
(Например, SAG1, LDH1, ENO1) в рН-сдвинуты попу-нормами в. В каждом из переходов развития, отобранных в данном исследовании была возможность обнаружить новую экспрессию генов паразита, и это остается область текущего расследования. Так, например, новый NTPase, которая связана с последовательностью с NTPI и II (Бермудес и др., 1994) был идентифицирован, экспрессия которого была обнаружена в тот же день 15-паразитов (но не на всех в день 7-паразитов) и был среди самых выраженных генов в рН-сдвинуты библиотеки (смотри раздел 16.3 ниже; Radke и белый, неопубликованные результаты). Комплексные списки генов, которые суммируют развитие конкретного выражения при каждом переходе можно найти на TgSAGEDB (https://vmbmod10.msu.montana.edu/ VMB / белой-лаборатория / newsage.htm).
Предыдущие исследования спорозоидных инфекций показали, что дифференцировка активный и быстрый процесс как в пробирке и в естественных условиях (Иероним и др., 1998). Хотя относительно небольшое число меток-ERS наблюдал в этих экспериментах, результаты привели к предположению, что дифференциация от спорозоитов на стадию тахизоит была Эссен-ственно полным в первых 1-2 дней после спорозоитов инфекции. Измерения цельноклеточного мудрец рассказывает другую историю, так как мРНК пулы паразитов появляются даже 4 дня после спорозоитов инфекция сохраняет значительную часть спорозоитов специфического мРНКа, такого, что корреляция между этими бассейнами высока (спорозоиты против 4-я дня, рçíàê ðàâíî0,728). Это наблюдение было неожиданным и свидетельствует о том, что, пытаясь понять Toxoplasma инфекции, инициированный ооцист, спорозоиты конкретной биология может иметь существенное влияние на ранних стадии развития в промежуточном хозяине.
Глобальные сравнения SAGE наборов данных из типов I, II, и III лабораторных штаммов в контексте развития VEG при условии, другие Юнекс-pected результатов, которые освещают важные биологические различия между штаммами. Была четкая корреляция между активируемыми тегами SAGE, полученными из Вега Day-6 после спорозоитов населения и те, кто в типе деформации I-RH, которые не наблюдались по сравнению с другими популяциями первичными Векли или тип деформации. Этот уникальный РЕЛА-tionship в экспрессии генов может отражать общую биологию: население Дня 6 Век, как паразиты RH, отсутствие каких-либо доказательств спорозоитов или брадизоитным
Экспрессия мРНК, и растут со временем аналогично быстрым удвоением (. Иероним и др, 1998; Radke и др., 2001). Важно отметить, что SAGE библиотеки построены из типа II-Me49B7 и типа III-VEGmsj пара-сайты не подняли день-6 SAGE тегов, но в отличии от RH / Day-6 наборов данных, SAGE теги корреспондентского-диньте в брадизоитных генах были обнаружены в библиотеках из этих штаммов. Эта разница, вероятно, связано с большей мощностью VEGmsj и Me49B7 паразитов, чтобы войти в брадизоитный развитие-мента пути, где RH является развитием некомпетентным (Dubey и др., 1999). Это неизвестно, будет ли также применяется этот дифференциал структура ген Expres-Сьон к другому вирулентному типу I. штаммов которого Неэффективность при формировании кист ткани также хорошо известна (Сибла и Бутройд, 1992; Dubey, 1997, 1980), но отсутствие гена брадизоитных затравок в вирулентных штаммах, что очевидно при анализе микрочип авирулентных штаммов недавно сообщалось (Saeij и др., 2005). Таким образом, структуры мРНК, обнаруженные в лабораторных штаммов по-видимому, отражают характеристики от естественного пути развития с их сравнительной позиции по отношению к росту смещенной попу-нормами в которые происходят на 7-й день после спорозоитов Infec-ции (Иероним и др., 1998). Штаммы, чьи экспрессии генов помещают их влево (и ранее) эту границы развития, как RH, дополнительно удалены из брадизоитных дифференциаций, и могут быть более вирулентными. Напротив, экспрессия генов структура, согласующаяся с популяциями вправо (то есть свидетельство базальной экспрессии генов брадизоитных) указует на большую емкость для брадизоитных разработок-ления, который связан с авирулентностью (Saeij и др., 2005). Сортировка экспрессия генов Мех-низмы, которые управляют этими переходами развития и понимание того, как изменения в этих элементах приводят к изменению деформационного потенциала развития является важной областью для будущих исследований.
Транскрипционные
УПРАВЛЕНИЕ В Toxoplasma
Синтез РНК в Toxoplasma
Базальный транскрипционный комплекс, который контролирует экспрессию кодирующих белок генов (класс II),
| Транскрипционные CONTROL IN Toxoplasma |
в большинстве эукариот РНК-полимеразы II, и связанные с ним общие факторы транскрипции (GTFS). Сравнение этих факторов транскрипции, а также сходство в трех ядерных полимеразах (Ranish и Hahn, 1996), показало, что эти механизмы в значительной степени консервативные в процессе эволюции, от Archea до млекопитающих. В хорошо изученных одноклеточных и многоклеточных эукариот, транскрипции включает в себя ряд взаимодействующих регуляторная комплексов, которые работают совместно, чтобы контролировать синтез РНК из определенной области генома. Активирование факторы транскрипции (АССАЛЙ) связывается с цис-действующим промотор элемента (ов) и рекрутировать хроматин ферментов, которые расслабляют хроматин вокруг циса-элемент, содержащей области, а также набора Mediator комплекс мульти-субъединицы, которая контактирует с РНК-поли-merase II, предварительно инициация комплекс (ПИК) непосредственно (Блазек и др., 2005). Доступность цис-элемента на связывание АТФ зависит от взаимодействия с этими ремоделирования ферментов, но также может быть под влиянием других факторов, таких как хроматина состояния в регуляторной последовательности и фазы клеточного цикла (Фрай и Петерсон, 2002). В своей очереди, расслаблено хроматин состояние позволяет для формирования ПОС в основе промоторов элементов через деятельность, содержащуюся в Mediator, которые облегчают набор РНК поли-merase II и GTFs. Следует отметить, что существующие модели изменения вида трансмиссионными жидкостями, чтобы предположить, что активация РНК-полимеразы II этими факторами происходит косвенно, через их вербовке АТФ-зависимых ремоделирования хроматина комплексов (Фитэрстоун., 2002; YJ Ли и др, 2004; Блазек и др., 2005). Кооперативная модель, описанная здесь, является общим, хотя вариация этой базовой модели в настоящее время хорошо известна в исследованиях отдельных генов (для обзора см Фрай и Петерсона, 2002).
В каких-то простейших есть примеры различий в регуляции генов, кодирующих белок. Например, в то время как РНК-полимеразы II-опосредованной транскрипции генов класса II в трипаносом было документально подтверждено, транскрипция VSG и проциклической кислой повторяющиеся белок (PARP) генов, как было показано, зависит от РНК-полимеразы I (Белли, 2000). Это необычно для эукариот, как это полицистронная
природа трипаносом транскриптов и пост-транскрипционной регуляции экспрессии генов (Belli, 2000). Анализ генов, кодирующих белки в Apicomplexa указует на то, что CONVEN-ционный РНК-полимераза с сходством с другим крауном эукариота присутствует (недавно рассмотрены в Мейснере и Soldati, 2005). Гомологи для всех известных требуемых эукариотических РНК-полимераз были обнаружены в геноме Toxoplasma: РНК-полимеразы I (транскрипция рибосомальной РНК), РНК-полимеразы II (расшифровывает белок-кодирующие транскрипты), и РНК-полимеразы III (расшифровывает малые РНК) (Ли и др, 1989, 1991,.. Фокс и др, 1993; Мейснер и Soldati, 2005). Кроме того, эта классификация подтверждается исследованиями в Plasmodium, показывающие, что синтез 18S рРНКα-amanitin нечувствительным, в то время как смысловая и антисмысловая транскрипция генов класса II ингибируется этой полимераза II-специфический токсин (Милителло и др., 2005). Эти результаты указывают на то, что плазмодий и токсоплазмы обладают законсервированный эукариотической техники в результате чего РНК-полимеразы II, расшифровывает белок-кодирующие гены.
Основные элементы эукариотических промоторов класса II включают в себя TATA бокс, инициатор (INR), а также вниз по течению элементы промотора (DPE), которые распознаются и связанные несколько GTFs: TFIIA, TFIIB, TFIID, TFIIE, TFIIF и TFIIH (Ranish и Хан, 1996; Физерстоун, 2002; Блазек и др, 2005;.. Ruvalcaba-Салазар и др, 2005). Ядро семейства GTF включает в себя белок, связывающий TATA (ТБФ), TFIID и РНК-полимеразы II. Исследования белка Plasmodium ТБФ показали, что эта часть TFIID комплекса действительно связывается со специфическим коробками элемента TATA в промоторной области (Ruvalcaba-Салазар и др., 2005), хотя неспособность найти и другие основные элементы, привело к предположению, что обычный двудольный Mech-ханизм из цис- и базальной транскрипции либо отсутствует, либо более примитивный в этих паразитах (Коулсон и др., 2004; Каллебает и др., 2005; Мейснер и Soldati, 2005). (. Callebaut и др, 2005) Совсем недавно, сочетание двумерного гидрофобного кластерного анализа с профильными на основе методов поиска с использованием PSI-BLAST продлил ранее исследования (Коулсон и др., 2004; Meissner и Soldati, 2005), такие что ~ 60 процентов известных эукариотических GTFs в
426 GENE РЕГУЛИРОВАНИЕ
Plasmodium генома, в том числе важных компонентов TFIID комплекса, в настоящее время определены. Подобные «в силикомарганце» изыскания для коих-нентов эукариотического транскрипционного аппарата демонстрируют наличие этих факторов в геноме токсоплазмы. Гомологи для различных субъединиц для GTFs (TFIID, TFIIE, TFIIF и TFIIH) и субъединицы Mediator (mdt6) (Мейснер и Soldati, 2005; Behnke и белого, неопубликованного) были найдены, и сравнение с вновь обнаруженным GTFs Plasmodium (. Каллебаут и др, 2005) показывает, что гены со значительным сходством существуют также в Toxoplasma: TFIID-Taf1 к TGG_993790; TFIID-Taf2 TGG_993790; TFIIE-αсубъединица TGG_994351-TgTwinScan_5763; TFIIE-βсубъединица TGG_994289-TgSwinScan_3074; TFIIF-βсубъединица TGG_994699-TgTwinScan_3381; TFIIH-TFB5 TGG_994580. Следует также отметить, что компоненты CCR4-комплекс не были идентифицированы в данных экспрессии токсоплазмы SAGE, в том числе гомологичных последовательностей для CCR4 субъединицу 7, not1 и Caf-1 (Behnke и белый, неопубликованной). Этот комплекс был законсервирован от дрожжей до человека, и служит в качестве регулирующего элемента для реагирования на изменения питательных веществ на уровень и стресс через взаимодействие с ПОС комплексом (Collart, 2003). В целом, эти investi-дина показывает, что в то время как GTFs менее консервативные в Apicomplexa, большая части базального транскрипционного аппарата и хроматина REMOD-ELING факторы (смотрите раздел 16.4), необходимый для медно-ческого контроля транскрипции генов эукариотов присутствуют в этих патогенах,