ГЕНЕТИЧЕСКАЯ ИНЖЕНЕРИЯ РАСТЕНИЙ
В течение тысячелетий усилия стихийной или сознательной селекции были направлены на повышение количества и качества урожая, причем без знаний молекулярной основы этого процесса. Получение первых трансгенных растений, т. е. растений, в геноме которых работают чужеродные гены различного происхождения (гены бактерий, дрожжей, млекопитающих, человека, гены эволюционно далеких видов растений), свидетельствует о больших возможностях генной инженерии в области селекции. Методология генетической инженерии растений сложнее, чем методология генетической инженерии бактерий. Она стала развиваться значительно позднее. Первые эксперименты по переносу в растения чужеродных генов датируются 1980 г. Так, в 1983 г. получен каллус, а позднее — химерное растение санбин {sunflower — подсолнечник и bean — бобы), представляющее собой подсолнечник, в геноме которого работают гены запасного белка фасоли — фазеолина.
В конце 80-х гг. получены трансгенные растения, устойчивые к гербицидам и к атаке насекомых.
Задачи и проблемы генетической инженерии растений
Основная цель генетической инженерии растений — выделение гена, определяющего полезный для культуры признак, и перенесение его в геном другой культуры, где этот новый ген должен стать частью наследственного аппарата.
Молекулярные биологи обратили внимание на растения как на объект исследования относительно недавно. Одна из причин — отсутствие среди них такого модельного объекта, каким стала Е. coli. Сейчас реальным кандидатом на эту роль становится двудольное растение арабидопсис. Интерес, проявленный к нему молекулярными генетиками, объясняется очень коротким циклом его генерации, малым размером его генома и удобством для клонирования генов.
Работа с растениями имеет одно немаловажное преимущество — они могут быть регенерированы из клеточной массы (каллуса) или из
недифференцированных соматических тканей в зрелые, способные к размножению (фертильные) растения.
Однако на пути использования новой технологии гибридных ДНК возникает серьезное препятствие — это отсутствие основательных знаний о регуляции работы генов в процессе развития растения.
Понятие «генетическая инженерия растений» включает работы на клеточном уровне и учитывает все аспекты культуры клеток и тканей, молекулярную биологию и перенос генов. Основное препятствие для введения ДНК в растительные клетки — клеточная стенка. Поэтому используют растительные протопласты (отдельные растительные клетки, стенки которых удалены обработкой целлюлолитическими ферментами), аналогичные сферопластам дрожжей, которые в питательной среде сохраняют жизнеспособность и при определенных условиях культуры образуют новые клеточные стенки с последующим делением и регенерацией из них целых растений. Разработан метод слияния протопластов и получения на их основе соматических гибридов.
В настоящее время на первый план выдвигаются две проблемы. Первая — идентификация и выделение генов, предназначенных для переноса в растение с целью приобретения им нового полезного признака. Вторая — разработка простых и доступных методов этого переноса с последующей работой новых генов в растениях.
Методы переноса генов в растения можно разделить на две основные группы: 1) с помощью векторных систем и 2) альтернативные методы прямого переноса.
Несмотря на все перечисленные сложности, генетическая инженерия растений вносит два существенных изменения в селекционную программу. Первое — это потенциальная экономия места и времени, поскольку получение образцов в лабораторных условиях исключает необходимость использования больших площадей для выращивания тысяч культур и может сократить время их созревания. Второе — это введение элемента точности в процесс селекции, поскольку исследователь получает возможность манипулировать определенным материалом в виде уже известных нуклеотидных последовательностей.
Агробактериальные трансформирующие
Векторы растений
Агробактерии и ризобии относятся к семейству Rhizobiaceae. Бактерии этих родов способны взаимодействовать с высшими растениями, они вызывают различные новообразования. Так, после инфицирования культур агробактериями образуются опухоли, а штаммы ризобий — симбионты — вызывают образование корневых клубеньков, способных осуществлять азотфиксацию у бобовых.
С проблемами генетической инженерии растений непосредствен-
но связан процесс опухолеобразоваиия. В случае агробактерий образуются настоящие опухоли с неограниченным опухолевым ростом клеток хозяина. Вирулентные их штаммы вызывают три вида рака у растений: корончатый галл (Л. tumefaciens), бородатый корень (Л. rhisogenes), стеблевой галл (A. rubi). Опухоли, вызванные агро-бактериями, представляют собой типичные неоплазмы, т. е. злокачественные новообразования, которые могут автономно размножаться в отсутствие бактерий. Это пока единственный пример опухолей среди растений и животных, которые вызываются бактериями.
После заражения растения агробактериями опухоли, развивающиеся из одной или нескольких клеток, быстро разрастаются в крупные образования. Иногда па периферии таких наростов формируются листовидные структуры, подобия побегов или корешков. Круг хозяев агробактерий чрезвычайно широк: голосемянные, большинство двудольных, некоторые однодольные (лилейные). Естественную устойчивость имеют кукуруза, пшеница, рожь, овес, рис, сахарный тростник, все низшие растения. В целом опухоли не убивают растения, но существенно подавляют их общий рост, жизнеспособность, повышают восприимчивость к грибным инфекциям и стрессам. Однако ряд растений погибают (например, виноградная лоза).
Корончатый галл как болезнь растений описай еще Аристотелем. В 1947 г. А. Браун культивировал ткани галла на искусственной среде с сахарозой и неорганическими солями, они росли быстро даже без добавления гормонов (цитокииов и ауксинов). Оказалось, что для индукции опухоли достаточно лишь кратковременного контакта бактерий с растениями, а само ее развитие может происходить в отсутствие бактерий, т. е. после заражения размножение опухолевых клеток происходит независимо от их присутствия. Эти наблюдения позволили Брауну заключить, что растительные клетки каким-то образом трансформируются после воздействия агробактерий. На основании этого было выдвинуто предположение, что бактерии вводят в хозяйскую клетку некий фактор, приводящий к перерождению нормальных клеток в клетки корончатого галла. Этот фактор ответственен лишь за первичную индукцию опухоли, но не за ее последующее развитие. В результате его действия изменяются или полностью прекращаются нормальные процессы роста и дифференциации растительной ткани.
В дальнейших исследованиях выявилось, что по своей химической природе опухолеобразующий фактор представляет собой нуклеиновую кислоту. Исследователи стали определять фракции ДНК, присутствующие в онкогеиных штаммах и отсутствующие в неонко-генных. Бельгийские исследователи выявили, что онкогенность агробактерий обусловлена присутствием в них группы ковалентио замкнутых кольцевых плазмидных ДНК (1—2 копии в бактерии). Эти плазмиды получили обозначение Ti {tumor inducing).
В 1977 г. генетики из США доказали, что опухоли возникают в
результате включения определенного фрагмента Ti-плазмид агробак-терий в растительную ДНК. Он получил обозначение Т-ДНК (transfer — перенесенный), т. е. инфекционный процесс, вызванный агро-бактериями, развивается на генетическом уровне.
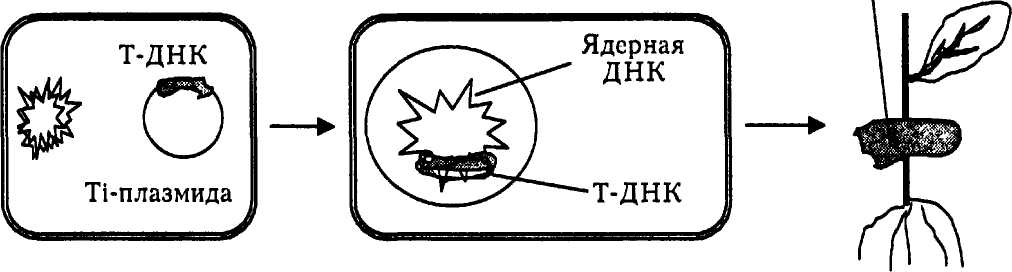
На основе экспериментальных данных можно представить следующую картину образования корончатых галлов (рис. 4).
|
Agrobacterium tumefaciens
Трансформированная растительная клетка
Корончатый
галл
Рис. 4. Схема образования корончатых галлов
Агробактерии, содержащие специфические Ti-плазмиды, проникая через поврежденную, раневую ткань, заражают растения. Часть плазмидиой ДНК попадает в растительную клетку. Т-ДНК вырезается из Ti-плазмиды и встраивается в хромосому растительной клетки. В результате фрагмент бактериальной ДНК становится частью наследственного аппарата растения и начинает экспрессироваться наряду с генами культуры. Его работа запускает процесс опухолеобразования.
Во-первых, начинается неконтролируемое деление клеток, в результате чего образуется опухоль. Во-вторых, в опухолевых клетках синтезируются необычные соединения, не встречающиеся больше нигде в природе, названные опинами; по ним можно тестировать опухолевую клетку.
Опины (производные обычных участников метаболизма — аминокислот и различных кетокислот или Сахаров) — биологически активные соединения нового класса, обнаруживаемые только в тканях корончатых галлов у растений. Эти низкомолекулярные метаболиты служат ростовыми веществами для агробактерии.
В-третьих, появляется еще одно новое свойство — гормононеза-висимый рост опухолевых растительных клеток.
Биологический смысл этого явления заключается в том, что Ti-плазмида действует как природный инструмент для переноса новых генов в растения, следовательно, может служить экспериментальным вектором для введения любых ДНК в геном растений. Молекулярных биологов интересует механизм переноса и встраивания Т-ДНК в растительную хромосому. Знание этих процессов оказалось очень важным для осуществления экспериментального введения чужеродных генов в растения.
Векторы на основе Ti-плазмид агробактерий. Обнаружение естественной системы переноса генов в растения с помощью Ti-плазмид стимулировало исследования в направлении использования их как векторов для генетической инженерии растений. Т-ДНК обладает свойствами, которые весьма желательны для универсального вектора генетической инженерии растений, а именно: широким спектром хозяев, эффективным способом инфекции, транскрипционной активностью и генетической стабильностью. Достаточно высока ее емкость: большие фрагменты чужеродной ДНК (длиной 50 тыс. пар нуклео-тидов и более), экспериментально встроенные в Т-ДНК, эффективно переносятся и внедряются в растительную хромосомную ДНК. Прежде чем природная Ti-плазмида будет использована как векторная система, ее необходимо модифицировать (рис. 5).
|
|
Х-ген
ДНК
Е. coli
A. tumefaciens
Т-ДНК
Х-ген
|
| Ядерная ДНК |
| Х-ген |
| Х-ген |
Т-ДНК
I

|
Трансгенное растение
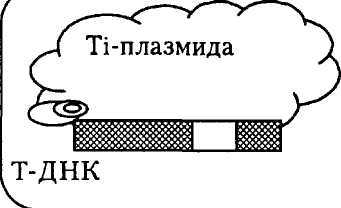
Рис. 5. Схема клонирования чужеродного гена в растении с использованием Ti-плазмид агробактерий
Для этого первоначально ген X, который нам необходимо ввести в растения, клонируют в клетках Е. coli с использованием соответствующих векторных систем. Далее его в составе подходящего вектора вводят в клетки Л. tumefaciens, где происходит встраивание гена X в Ti-плазмиду, а именно в Т-сегмент. Затем такой рекомбинантной бактерией обрабатывают клетки растений, где Т-ДНК, несущая ген X, встраивается в ядерную ДНК растительной клетки. В результате кло-
иирования таких рекомбинаитных растительных клеток с последующим регенерированием из них растений получаются трансгенные растения, в геноме которых работает чужеродный геи X (см. рис. 5).