Полисахариды (или гликаиы) — полимеры, построенные не менее чем из 11 моиосахаридных единиц одного или нескольких типов. Соответственно различают гомополисахариды и гетерополисахариды. Они являются обязательными компонентами всех организмов и составляют большую часть углеводов, встречающихся в природе, преобладающую долю в биомассе растений, а следовательно, и основную массу органического вещества на Земле.
Полисахариды встречаются в виде самостоятельных полимеров, а также в комплексах с нуклеиновыми кислотами, белками, липидами, фосфатом. Существует множество их видов, различных по мономер-пому составу и структуре. Особым разнообразием отличаются полисахариды микроорганизмов. Некоторые из них близки или идентичны гликанам растений и животных. Но подавляющее их большинство имеет уникальную структуру, специфичную для вида или для его серологической группы. В микробных гликаиах часто обнаруживаются ранее неизвестные моносахара, которые не встречаются ни у животных, ни у растений.
О том, что слизь, образуемая многими микроорганизмами, может иметь углеводную природу, знали еще во времена Пастера. Однако особое внимание микробным гликанам стали уделять лишь с начала 20-х гг. прошлого столетия, когда узнали, что вещества, определяющие серологическую специфичность пневмококков, являются полисахаридами. В настоящее время исследование этих полимеров приобрело особое значение в связи с открывшейся возможностью широкого их применения в медицине и ряде областей народного хозяйства.
Полисахариды микроорганизмов в зависимости от локализации делятся на внутриклеточные и внеклеточные. К первым относят обычно гликаиы цитоплазмы, мембран и клеточных стенок, а ко вторым — капсул, чехлов (пристеночные структуры) и свободной слизи, не прилегающей к клеточной стенке. Иногда среди внеклеточных встречаются полисахариды, локализованные вне цитоплазматической мембраны. В этом случае в группу внеклеточных попадают и гликаиы клеточных
стенок. У ряда микроорганизмов действительно трудно различить границу между капсулой и клеточной стенкой. Поэтому нередко полисахариды делят на три разновидности: внутриклеточные (цитоплазмы, мембран, периплазмы), клеточных стенок и внеклеточные (капсул, чехлов и свободной слизи).
Термин «экзогликаиы» применяют в основном к полисахаридам свободной слизи. Иногда так называют также капсульпые их разновидности.
Микробные полисахариды объединяют в группы и по функциям: резервные, участвующие в активном транспорте; опорные, участвующие во взаимодействии между клетками; защитные и др.
Внеклеточные полисахариды, капсульпые или свободные или те и другие, образуют многие микроорганизмы. Пожалуй, нет такой группы последних, представители которой не обладали бы этой способностью. Однако синтез внеклеточных полисахаридов — необязательная функция клетки, и проявляется она лишь в определенных условиях. Встречаются микроорганизмы, у которых не было замечено пи капсул, пи слизи.
Внеклеточные полисахариды микроорганизмов чрезвычайно разнообразны по составу и строению. К настоящему времени исследован состав около 200 экзогликанов, установлены первичная структура и детали строения многих из них. В составе внеклеточных полисахаридов различных микроорганизмов обнаружено более 20 моносахаридов и их производных. Наиболее часто встречаются гексозы: глюкоза, галактоза, манноза и 6-дезоксигексозы (например, рамноза). Реже выявляются пентозы: арабипоза, ксилоза, рибоза. Распространены уро-иовые кислоты: галактуроиовая, мапиуроновая и особенно глюкуро-иовая. Многие содержат амииосахара: глюкозамип, галактозамин и машюзамии. Часто в экзогликаиах присутствуют неуглеводиые заместители — пиру ват и ацетат, встречаются также сукцинат и глицерин. Для ряда внеклеточных гликанов характерно наличие редких мономеров: 2,6- или 3,6-дидезоксисахаров; у некоторых найдены ранее неизвестные моносахариды, например, гликолактиловые кислоты (у сапротрофиых микобактерий). Иногда обнаруживаются тейхоевые кислоты, фосфатные и сульфатные ионы.
Внеклеточные полисахариды большинства видов бактерий — кислые гетерогликаны разнообразного состава, построенные из 2 — 5, иногда 6 — 7 мономеров, линейные и разветвленные, имеющие регулярную структуру из повторяющихся олигосахаридиых звеньев. Так, например, Xanthomonas campestris синтезирует полиаииоиит ксаптап, включающий глюкозу, маннозу, глюкуроновую кислоту, О-ацетильную группу и пируват.
Некоторые бактерии образуют нейтральные гетерополисахариды.
Весьма распространены у микроорганизмов различные гомополи-сахариды, особенно глюкаиы, из которых наиболее известны декстра-иы (группа более или менее близких по строению нейтральных глю-
капов). Они могут содержать до 200 000 остатков глюкозы, бывают линейными и разветвленными. Линейная (основная) цепь построена при участии а-1,б-связей, ветвление обусловлено а-1,2-, а-1,3- и а-1,4-связями. Молекулярная масса декстраиов колеблется от 12 до 600 млн. Наиболее активные их продуценты — представители молоч-по-кислых бактерий Leuconostoc mesenteroides и L. dextranicum. Они синтезируются также некоторыми видами Streptococcus (Str. sanguis, Str. mutans), Brevibacterium, Lactobacillus. Практически каждый продуцент синтезирует свой, несколько отличный от других видов декстран. Некоторые штаммы образуют одновременно две его структурные разновидности.
Внеклеточную целлюлозу — полисахарид, распространенный в растениях, — из бактерий синтезируют некоторые представители Pseudomonas, Zooglea, Azotobacter, подавляющее число видов Rhizobium и Agrobacterium, Acetobacter xylinum. 0-(1-3)-Глюкап, называемый курдланом, образуют Alcaligenes faecalis var. myxogenes и виды Rhizobium. Сильно разветвленный а-(1->4)-глюкан с боковыми цепочками, присоединенными а-(1->б)-связями, — полимер типа гликогена, резервного полисахарида животных, многих бактерий и дрожжей, — можно обнаружить в культуралыюй жидкости Neisseria perflava. Нигеран — а-глюкан с чередующимися (1-4)- и (1-3)-связя-ми — образует гриб Aspergillus niger.
Разветвленные полимеры фруктозы — леваны с (2-6)-связями — синтезируют уксусно-кислые бактерии Gluconobacter oxydans, Acetobacter aceti, некоторые виды Pseudomonas, Erwinia (подгруппы herbicola), Aeromonas, Bacillus. Фруктаиы типа инулина (резервный полисахарид растений семейства сложноцветных) с (2-1)-связями в основной цепи и ответвлениями в положении С6 образуют штаммы Str. mutans.
Манианы обнаружены в культурах некоторых видов Bacillus и Corynebacterium, а также у многих дрожжей.
Чаще всего микроорганизмы, способные к образованию внеклеточных полисахаридов, синтезируют капсулы и свободную слизь. Мономериый состав последних в большинстве случаев одинаковый.
Не всегда можно определить, какие именно полисахариды свойственны той или иной группе микроорганизмов. Часто филогенетически близкие бактерии синтезируют внеклеточные гликаны, сходные по составу и строению. К ним относятся, например, бактериальный альгинат Pseudomonas aeruginosa и Azotobacier vinelandii, курдлап Rhizobium и A. faecalis var. myxogenes, кислые гетерогликаны Corynebacterium и Arthrobater, декстраны Streptococcus и Lactobacillus и др. Однако нередко микроорганизмы, далеко отстоящие в таксономическом отношении, образуют гликаны сходного состава или одинаковые. Обычно в этом случае полимеры проявляют и функциональное сходство. Так, очень близки по составу экзогликаны различных фито-патогенных бактерий и возбудителей менингита. У ряда бактерий це-
ментирующим материалом при образовании клеточных агрегатов служит внеклеточная целлюлоза.
Известно, что не только разные виды одного рода микроорганизмов, но часто и разные штаммы одного вида синтезируют неодинаковые экзополисахариды. Так, Е. coli имеет около 70 серотипов, различных по составу и структуре капсульиых полисахаридов и, следовательно, по иммуиохимическим свойствам. Экзогликаны эффективных и неэффективных штаммов клубеньковых бактерий различаются по моио-мериому составу, а разных штаммов дрожжей рода Lipomyces — по соотношению моносахаров. Неодинаковыми по моиосахаридиому составу могут быть внеклеточные гликаиы М-, S- и #-форм бактерий.
Отмечены случаи, когда в культуралыюй жидкости одного микроорганизма накапливается несколько различных гликаиов. Например, L. mesenteroides образует декстраи и леван, у Serratia marcescens обнаружено три экзополисахарида: кислый глюкорамиан, содержащий глюкуроиовую кислоту, рамиоглюкаи и гептоглюкаи. Гриб Aureobasi-dium (Pullularia) pullulans образует два гомоглюкаиа: аубазидаи — разветвленный полимер с а-(1-4)-, (3-(1-3)- и Р-(1-б)-связями, причем в боковой цепи может быть от одного до четырех остатков глюкозы на одно звено триозы, и пуллулан — линейный глюкаи, состоящий из мальтотриозных и мальтотетраозиых фрагментов, соединенных а-(1-б)-связями.
Внеклеточные полисахариды не являются жизненно необходимыми для микроорганизмов. В природе есть виды, никогда их не образующие. Экспериментально показано, что клетки, лишенные капсул, столь же жизнеспособны, как и капсулировапиые. Тем не менее внеклеточные полисахариды выполняют определенные функции, способствующие поддержанию условий, благоприятных для жизнедеятельности продуцента. Одни из них универсальны для всех полисахаридов, поскольку определяются общими для этих веществ свойствами, другие специфичны для определенного гликана, что обусловлено особенностями состава и строения данного полимера.
Внеклеточные полисахариды предохраняют клетки от высушивания, от губительного действия ультрафиолетовых лучей и различных химических агентов, в том числе тяжелых металлов и лекарственных препаратов. Замечено, что капсулированпые бактерии устойчивее к химиотерапевтическим средствам, чем бескапсульиые варианты. Располагаясь поверхностно, капсулы защищают клетки от бактериофагов и поглощения их простейшими, предупреждают денатурацию клеточного белка.
Многие внеклеточные гликаны биологически активны и определяют иммунологические свойства и вирулентность штаммов. Как правило, чем толще капсула, тем выше вирулентность и патогенпость бактерий. Некоторые патогенные бактерии, лишенные капсул, становятся авиру-лентными. R-антигены являются агрессинами, подавляющими фагоцитоз бактерий и создающими таким образом условия для их размножения.
Одно из характерных проявлений биологической активности полисахаридов — способность модифицировать ферменты различных организмов. В результате стимулируется или снижается активность последних. Экзополисахариды фитопатогепиых бактерий являются фитотоксииами, участвующими в специфическом взаимодействии бактерий с растительной тканью. К ним относится, например, ксантан.
В некоторых случаях внеклеточные гликаиы служат резервным источником углерода и энергии, а азотсодержащие полисахариды — источником азота для продуцента. Полиаиионные полисахариды концентрируют катионы из окружающей среды и способствуют их транспортировке в клетку. Полисахаридио-липидиые комплексы микроорганизмов, растущих на средах с w-алканами (псевдомопад, иокардий, корииебактерий, дрожжей), — хорошие эмульгаторы. Они снижают поверхностное натяжение, увеличивают площадь соприкосновения углеводорода и воды, образуют мицеллы и тем самым способствуют проникновению углеводородов в клетку.
Нейтральные внеклеточные полисахариды поддерживают целостность нитчатых форм и различных скоплений клеток. Гликаиы, несущие определенный заряд, напротив, способствуют диспергированию клеток. Экзогликаны некоторых бактерий ответственны за прикрепление клеток к поверхности. Так, Str. mutans, вызывающий кариес зубов, прикрепляется к зубной эмали с помощью декстрана.
Биосинтез полисахаридов
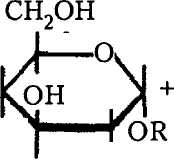
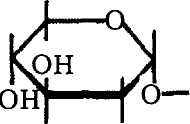
Биосинтез гликанов сводится к созданию гликозидной связи между моиосахарами. В общем виде это можно представить так: глико-зильный донор передает гликозил на акцептор-затравку, а сам высвобождается. Полимеризация идет вплоть до образования готового полисахарида. Процесс катализируется специфическими гликозилтранс-феразами, а ветвление полимеров — другими, «ветвящимися», ферментами, отщепляющими фрагменты линейной цепи недостроенного полисахарида и переносящими их на ту же или аналогичную цепь в определенное положение (рис. 8).
| ОН |
 а
а
ОН
сн2он
он
| он | |||
| Фермент | -л | ||
| .у | |||
| О | |||
| п | ОН |
сн2он
п
ROH
Рис. 8. Схема образования гликозидной связи: а — донор, б — акцептор
Биосинтез гетерополисахаридов проходит в двух вариантах: либо регулируемое чередование различных мономерных единиц, либо пред-
варительный синтез ди- и олигосахаридов, которые остаются присоединенными к донору, а затем полимеризуются.
Акцепторами при биосинтезе полисахаридов выступают олигоса-хара и недостроенные гликаиы, причем первые из них оказываются, как правило, первичными. Это наблюдается, например, в случае синтеза декстрапов и леванов (сахароза), целлюлозы (целлодекстрииы), хитина (хитодекстрины). Иногда первичным акцептором может быть недостроенный полисахарид, или затравка.
Доноры моиосахаридиого остатка имеют различную природу. Редко ими выступают олигосахара. Так, при биосинтезе декстранов и лева-пов донором может быть сахароза. Но в большинстве случаев сахара-доиоры должны находиться в активированной форме. Такое состояние моиомерпых единиц достигается при помощи иуклеозидтрифос-фатов с образованием нуклеозиддифосфатсахаров (НДФС), которые признаны наиболее распространенными и универсальными донорами гликозильпых остатков. Так, в биосинтезе целлюлозы Acetobacter xylinum участвует уридиидифосфатглюкоза, хитина Neurospora cras-sa — уридипдифосфат-М-ацетилглюкозамин, маииаиа дрожжей — гуаиозиидифосфатманноза. Очевидно, что в биосинтезе гетерополиса-харидов принимает участие несколько доноров соответствующих гликозильпых остатков. Взаимопревращения моносахаридов происходят на уровне НДФС. Они осуществляются специальными ферментами. В случае их нехватки одно из звеньев биосинтеза полисахарида блокируется; образуется дефектный полимер, или он не образуется вовсе. НДФС могут передавать не только моносахаридные, но, по-видимому, и олигосахаридные остатки.
Обязательными компонентами цепи биосинтеза полисахаридов у многих микроорганизмов являются наряду с НДФС и другие активированные сахарные интермедиаты — гликозилнесущие липиды. По строению это полиизопренолфосфатсахара. У большинства прокариот преобладают полипренольные носители с И изопрепоидными остатками — ундекапренолы (С55)- У микобактерий имеются переносчики с 10 остатками (С™)- Полипренолы эукариот более высокомолекулярны. Например, Sacch. cerevisiae содержит 16 — 18, a Asp. niger — 19 — 21 изопреноидных остатков. В микроорганизмах имеются полипреполмонофосфатсахара и полипренолпирофосфатсаха-ра, причем последних больше.
Количество гликозилиесущих липидов составляет всего десятые доли процента от массы клетки. Однако роль их важна и многообразна. Они могут быть промежуточными переносчиками моно- или оли-госахаридпых остатков, принимая их от НДФС и передавая акцептору — строящемуся полисахариду. Полипренолмонофосфатсахара присоединяют и переносят, как правило, только моносахаридный остаток, а полипреиолпирофосфатсахара — ди-, три- и тетрасахаридные звенья. В некоторых случаях перенос моно- и олигосахаридных остатков к акцептору происходит с изменением конфигурации глико-
зидной связи. На полипренолпирофосфатах может начинаться первичная сборка полисахаридной цепи. Первая стадия этого синтеза — построение олигосахарида с участием НДФС, затем происходит полимеризация компонентов первого, из которых начальный сохраняет связь с липидом, а остальные передаются акцептору. Гликозилиесущие ли-пиды участвуют и в транспорте Сахаров на внешнюю сторону мембраны и в клеточную стенку. НДФС не способны проходить через мембраны.
Считают, что биосинтез полисахаридов во многих случаях может осуществляться только за счет НДФС, но в реакциях, идущих на поверхности раздела между водной и липидиой фазой мембраны, необходимо участие гликозилнесущих липидов.
Предполагается, что остатки органических кислот включаются в молекулу полисахарида в соответствующей кофермептиой форме. Однако есть мнение, что в гликан может встраиваться только пиру-ват, который уже в молекуле полимера модифицируется в остаток определенной кислоты.
Вопрос о месте биосинтеза ряда полисахаридов, особенно внеклеточных, да конца не решен. Этот процесс у многих гликанов связан с определенными структурами клетки, так как соответствующие ферменты, НДФС и гликозилнесущие липиды локализованы в области цитоплазматической мембраны. Обнаруживаются они и во фракциях клеточных стенок. Однако существуют и внеклеточные ферментные полисахаридсинтезирующие системы. Это декстран- и левансахаразы большинства микроорганизмов, образующих соответствующие полисахариды. Хотя указанные ферменты не всегда целиком выделяются из клетки (у Gluconobacter oxydans в культуралыюй жидкости обнаруживается около 80 % левапсахаразы), сборка декстранов и леванов в значительной степени идет вне клетки. Но синтез большинства известных экзополисахаридов имеет внутриклеточную локализацию. Для экзогетерогликанов внеклеточный синтез вообще неизвестен. О том, в каком участке клетки происходит сборка полимера и как он выводится наружу, имеется несколько предположений. Согласно одному из них полимеризация гликана может завершаться на внешней стороне цитоплазматической мембраны, а выделение его вовне, вероятно, происходит через поры клеточных стенок. По другой гипотезе полисахарид образуется внутри находящихся в цитоплазме мембранных пузырьков, а затем с их помощью выносится из клетки обратным пино-цитозом. Если соответствующие ферменты локализованы на периферии клетки, то внеклеточные полисахариды иногда рассматриваются как результат сверхсинтеза гликанов клеточной стенки. Очевидно, что в этом случае полисахариды клеточной стенки и внеклеточные должны быть идентичны. Однако наблюдается это редко.
Полисахариды капсул и свободной слизи, одинаковые по составу и структуре, имеют, по-видимому, общее происхождение. Полагают, что при этом слизь образуется в результате отторжения капсульного материала.
in
Модель синтеза экзополисахаридов у бактерий, основанная на изучении этого процесса у Clebsiella aerogenes и родственных видов, представлена на рис. 9.
Локализация
Функция
Реакция
Внутри-клсточно и на мембране
Образование
предшественников
Глю------ ►Глю-б-Ф------ ►ГлкИ-Ф------- ►УДФ-Глю------- »УДФ
Гексокиназа Фогфоглюко- УДФ-глюко- I I Эпимераза I
мутаза пирофосфо- | | |
рилаза
I
I I I
| группа ферментов - - |
— — -I 1—П группа— -I ферментов
| УДФ — Глю 2УДФ — Гал УДФ - Глюкуроновая кислота |
Мембрана
Сборка повторяющихся
единиц
УМФ-у ГНЛ-Ф-
I
УДФ УДФ
Трансмутазы
удфн
Ф—Глю—Гал—Гал—Глюку роноваякислотаi
1------------------------------ III группа ферментов------------------------ '
Мембрана
Мембрана и клеточная стенка
Полимеризация
Перенос
на акцептор
ГНЛ — Ф — Ф — Глю — Гал — Гал — Глюкуроновая кислота+ +ГНЛ — ф — ф — Глю — Гал — Гал — Глюкуроновая кислота
I ГН Л1- Ф - Ф - Глю - Гал - Гал - Глю - Гал - Гал+ГНЛ - Ф1- Ф
1 II1
I Глюкуроновая Глюкуроновая '
1 кислота кислота '
L--------------- IV группа ферментов, полимеразы- - - -1
Акцептор - [ - Глю - Гал - Гал - ] + ГЛ Н - Ф - Ф
Глюкуроновая кислота
Примечание. Глю — глюкоза, Гал — галактоза, Глю-6-Ф — глююмо-6-фосфат, УМФ — уридинмонофосфат, УДФ — уридиндифосфат, ГНЛ — гликшилнесущий
липид
Рис. 9. Модель синтеза экзополисахаридов у бактерий
Влияние аэрации и температуры на биосинтез полисахаридов очень многообразно. Их режимы, благоприятные для образования того или иного гликана, могут сильно различаться.
Большинство микроорганизмов, образующих экзогликаны, — аэробы или факультативные анаэробы, поэтому в условиях хорошей аэрации выход экзогликанов в их культурах выше. Однако избыточное аэрирование может угнетать их биосинтез вследствие быстрого окисления углеродного субстрата.
В отношении действия температуры также наблюдается определенная закономерность. Максимальное образование гликанов часто
происходит при температуре ниже оптимальной для роста микроорганизмов. Так, количество декстрана в культуре L. mesenteroides увеличивается с уменьшением температуры с 30 до 10 °С. Но в некоторых случаях температурные оптимумы роста бактерий и образования ими экзогликанов совпадают.
Изменение темлературы культивирования микроорганизмов иногда приводит к синтезу полисахаридов с измененными свойствами. Например, известен штамм стрептококков, синтезирующий при 22 °С внеклеточный полисахарид, отличающийся более высоким содержанием глюкозамина и иными иммунохимическими свойствами, чем гли-кан, образуемый при 37 °С.