Посттранскрипционные модификации предшественников эукариотических мРНК по сравнению с теми же изменениями первичных транскриптов прокариот более разнообразны и играют большую роль в регуляции экспрессии их генов. Почти все эти реакции происходят в ядре эукариотических клеток в процессе синтеза РНК или сразу же после его завершения. Прежде всего, к 5’-концевому нуклеотиду большинства пре-мРНК присоединяются кэп-группы, что сопровождается метилированием одного или нескольких концевых нуклеотидов, в большинстве случаев необходимым для стабилизации и экспорта соответствующих мРНК из ядра в цитоплазму, а также их эффективной трансляции рибосомами. В основном те же функции, по-видимому, выполняет и полиаденилирование 3’-концевых последовательностей мРНК, которые подготавливаются к этому этапу путем специфического отщепления избыточных 3’-концевых нуклеотидов предшественника. Кроме того, интроны, содержащиеся в гигантских первичных предшественниках мРНК, с высокой точностью удаляются в результате сплайсинга. Ниже будут рассмотрены механизмы перечисленных посттранскрипционных модификаций пре-мРНК эукариот. Об использовании этих реакций в регуляции экспрессии генов на посттранскрипционном уровне речь пойдет в разделе 3.3.
Кэпирование. Сразу же после инициации транскрипции чаще всего происходит котранскрипционная модификация 5’-конца мРНК, сопровождаемая присоединением так называемой кэп-группы и дальнейшими ее изменениями. Кэпирование является одной из самых ранних модификаций растущих цепей РНК и происходит после полимеризации ее первых 20–30 нуклеотидов. Такая котранскрипционная модификация мРНК не только стабилизирует мРНК в
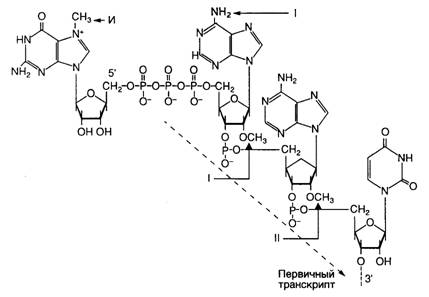
Рис. I.12. Обобщенная структура кэп-группы эукариотических мРНК
Указаны сайты метилирования, характерные для кэп-групп разных типов. И – инвариантный сайт, метилируемый у кэп-групп всех типов, I, II – сайты, метилируемые у кэп-групп типов 1 и 2 соответственно
цитоплазме, но и необходима в большинстве случаев для ее эффективной трансляции. Так, один из факторов инициации трансляции eIF-4E выполняет функции кэп-связывающего белка и требуется для осуществления кэп-зависимой трансляции мРНК. Кроме того, установлено, что кэпирование мРНК необходимо для эффективного сплайсинга пре-мРНК, ее полиаденилирования и экспорта из ядра в цитоплазму. Кэпированию подвергаются только транскрипты РНК-полимеразы II. На исключительную значимость реакций кэпирования указывает и тот факт, что контролирующие их гены являются жизненно важными.
Как уже упоминалось выше, транскрипция у эукариот и прокариот начинается, как правило, с пуринового рибонуклеозидтрифосфата – ATP, или GTP, причем трифосфатная группа сохраняется в составе мРНК. Таким образом, 5’-концевая последовательность мРНК в ядре на ранних этапах транскрипции представлена в следующем виде: ppp(A/G)pNpNpN... Гуанилилтрансфераза катализирует присоединение к растущей цепи мРНК молекулы GMP, которая оказывается связанной с 5’-концевым пурином 5’–5’-трифосфатной группой. Суммарная реакция первого этапа процесса кэпирования выглядит следующим образом:
G (5’) ppp + ppp(5’)(A/G)pNpNpN...
¯ Гуанилилтрансфераза
G (5’) p pp(5’)(A/G)pNpNpN... + pp + p
Реакция, по-видимому, протекает в две стадии. Вначале фермент связывает молекулу GTP (входящую затем в состав кэп-группы), что сопровождается отщеплением пирофосфата и образованием ковалентной связи фермент–GMP. Далее GMP присоединяется к 5’-концу мРНК, которая в результате теряет g-фосфатную группу. В результате нуклеотид кэп-группы оказывается в обратной ориентации по отношению к остальным нуклеотидам мРНК. Процесс создания кэп-группы этой последовательностью реакций не заканчивается. На заключительных этапах кэпирования происходит метилирование по N7 ранее присоединенной молекулы гуанозина. Такие посттранскрипционные модификации происходят в несколько стадий в цитоплазме клеток после транспорта процессированной мРНК из ядра с участием цитоплазматических ферментов.
Первая стадия метилирования осуществляется ферментом РНК(гуанил-7)-метилтрансферазой, которая переносит метильную группу S-аденозилметионина в положение 7 концевого гуанина кэп-группы (рис. I.12). Кэп-группа, метилированная лишь по этому положению, характерна для одноклеточных эукариот и получила название кэпа 0-го типа. Вслед за этим у большинства многоклеточных эукариот происходит метилирование 2’-ОН рибозы 5’-концевого инициаторного нуклеотида (A или G), который является первым нуклеотидом, включаемым в мРНК при инициации ее синтеза РНК-полимеразой. Метилирование катализирует другой цитоплазматический фермент – 2’-О-метилтрансфераза. Такая основная форма кэпа большинства эукариот получила название кэпа 1-го типа. Очень редко и только у тех мРНК, инициация синтеза которых происходит с ATP, под действием 2’-О-метиладенозин-N6-трансферазы метилируются NH2-группы этого остатка А. Фермент распознает данную концевую группу в качестве субстрата лишь в том случае, если она была предварительно метилирована в положении 2’-OH в результате вышеописанной реакции.
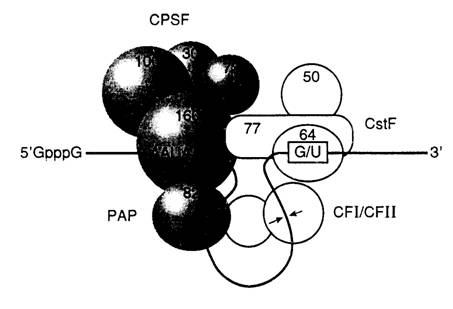
Рис. I.13. Модель процессинга 3¢-концевых последовательностей эукариотических пре-мРНК
Цифрами указаны молекулярные массы белков, входящих в состав процессирующего комплекса. Обозначены кэп-группа, последовательность поли(А)-сайта и регуляторная последовательность, с которой взаимодействует гетеродимерный фактор, стимулирующий расщепление (CstF). Стрелки указывают место расщепления РНК
PAP – поли(А)-полимераза, CPSF – фактор специфичности и расщепления РНК, CF I и CF II – факторы расщепления I и II
У некоторых видов эукариот метильная группа может дополнительно присоединяться ко второму от кэп-нуклеотида нуклеозиду мРНК (см. рис. I.12). Субстратом для этого фермента служит мРНК с кэпом 1-го типа, уже содержащим две метильные группы. В результате происходит метилирование остатка рибозы по 2’-ОН-группе с образованием структуры, получившей название кэпа 2-го типа. Если эта реакция имеет место, то мРНК, содержащие кэп 2-го типа, составляют 10–15% от общей популяции молекул кэпированных мРНК.
Иная структура кэп-группы характерна для некоторых зрелых некодирующих РНК, в частности малых ядерных РНК, обогащенных урацилом (U-мяРНК). В этом случае остаток гуанозина кэп-группы дважды метилирован в положении 2 в дополнение к обычной метильной группе в положении 7: m2,2,7G(5')ppp(5')N. Такое гиперметилирование U-мяРНК требуется для импорта собранных U-мяРНП-частиц в ядро и, возможно, предотвращает вовлечение U-мяРНК в трансляцию.
Полиаденилирование. Одним из обязательных этапов созревания предшественников эукариотических мРНК, синтезированных в ядре, является процессинг их 3’-концевых последовательностей, тесно сопряженный с присоединением кэп-группы. Созревание 3'-конца мРНК является двухэтапным процессом. Вначале предшественник теряет 3’-концевую некодирующую последовательность, после чего, как правило, к 3’-концу присоединяется поли(А)-последовательность путем ферментативной полимеризации остатков AMP:
5’ GpppG__________AAUAAA__¯________UUUUU___ 3’
¯ Расщепление
5’ GpppG__________AAUAAA__OH P____UUUUU___ 3’
¯ Полиаденилирование
5’ GpppG__________AAUAAA__AAAAAAAAAAAAAAA 3’
В настоящее время известно несколько исключений из этого правила: гистоновые мРНК животных и мРНК некоторых вирусов, предшественники которых расщепляются с помощью высокоспецифических эндонуклеаз и не полиаденилируются. Остаются неполиаденилированными и U-мяРНК, которые также являются транскриптами РНК-полимеразы II. В этом случае кэпированный первичный транскрипт мяРНК U1, содержащий на своем 3'-конце несколько избыточных нуклеотидов, экспортируется из ядра в цитоплазму, где и происходит удаление избыточной последовательности, которое в ядре блокировано специфическим белковым ингибитором TPI (3'-terminal processing inhibitor).

Рис. I.14. Модель белкового комплекса, элонгирующего поли(А)
PAP – поли(А)-полимераза, PAB II – поли(А)-связывающий белок II
Места отщепления 3’-концевых некодирующих последовательностей в мРНК животных обычно маркированы специальными последовательностями нуклеотидов (рис. I.13). Имеются, по крайней мере, две такие последовательности, образующие сайты полиаденилирования, или поли(А)-сайты. Одна из них – AAUAAA расположена за 15 нуклеотидов перед расщепляемой фосфодиэфирной связью и практически одинакова у всех исследованных организмов. Другая, менее изученная последовательность располагается сразу же за первой и часто состоит из нескольких остатков U или обогащена GU. Сайт расщепления РНК определяется расстоянием между этими двумя элементами с предпочтительным расщеплением фосфодиэфирной связи на 3’-конце остатка A, находящегося на участке, в котором расщепление разрешено. Имеются данные о том, что последовательности, расположенные выше AAUAAA, могут оказывать стимулирующее влияние на процессинг, но их присутствие необязательно для его правильного осуществления.
С последовательностью AAUAAA взаимодействует фактор CPSF (cleavage and polyadenylation specificity factor), определяющий специфичность расщепления и полиаденилирования РНК. CPSF состоит из четырех субъединиц с молекулярными массами 160, 100, 70 и 30 кДа. Последняя из них, по-видимому, не является необходимой для его функционирования. Самая большая субъединица находится в непосредственном контакте с последовательностью AAUAAA.
С GU-богатой и ниже расположенной последовательностью связывается гетеродимерный белковый фактор CstF (cleavage stimulating factor), стимулирующий расщепление и состоящий из трех субъединиц (77, 64 и 50 кДа). Вторая субъединица контактирует с GU-богатым регуляторным элементом и обладает типичным РНК-связывающим доменом. По отдельности факторы CPSF и CstF лишь слабо взаимодействуют с РНК. Однако их одновременное присутствие приводит к образованию прочного комплекса. Такой кооперативный эффект и взаимодействие двух факторов между собой определяются их большими субъединицами.
В расщеплении РНК непосредственно участвуют еще два фактора: CFI и CFII (cleavage factors). Как и в предыдущем случае, лишь вместе они образуют прочный комплекс с РНК.
Для полного реконструирования бесклеточной системы, осуществляющей процессинг 3’-концов in vitro, в ней помимо вышеупомянутых факторов необходимо наличие поли(А)-полимеразы – фермента, непосредственно осуществляющего полиаденилирование. Присутствие этого фермента требуется не для самого акта расщепления РНК, а, по-видимому, для стабилизации процессирующего белкового комплекса, схематически изображенного на рис. I.13. Сборка такого сложного комплекса зависит от ATP, однако в процессе сборки не происходит расщепления ее b–g-связей. В настоящее время неизвестно, какой именно компонент этого комплекса непосредственно расщепляет фосфодиэфирные связи РНК. Процесс полиаденилирования начинается сразу же за расщеплением РНК и происходит настолько быстро, что неполиаденилированных промежуточных продуктов не обнаруживается. Такое сопряжение двух реакций необходимо для защиты 3’-концевых последовательностей РНК от деградации нуклеазами. При этом сам акт полиаденилирования требует наличия только фактора CPSF, но не трех других: CstF, CFI и CFII.
Поли(A)-полимераза животных состоит из двух субъединиц с молекулярными массами ~ 80 и ~ 43 кДа, которые образуются в результате альтернативного сплайсинга их общей пре-мРНК. Короткий полипептид не обладает ферментативной активностью, и его функции неизвестны. Большая полипептидная цепь содержит С-концевой домен, обогащенный Ser и Thr и не определяющий ни одну из функций фермента, обнаруживаемых in vitro. Предполагают, что регуляторную роль играет множественное фосфорилирование этого домена. С-Концевой домен также содержит один из двух сигнальных последовательностей, необходимых для транспорта фермента в ядро. Вторая сигнальная аминокислотная последовательность локализована на границе С-концевого домена и основного полипептида поли(A)-полимеразы. Сравнительное исследование первичной структуры поли(А)-полимеразы показало наличие в ее полипептидной цепи каталитического домена, характерного для полимераз так называемого семейства X, к которому относятся ДНК-полимераза b, терминальная трансфераза, а также некоторые другие нуклеотидилтрансферазы. Используя 3’-конец расщепленной РНК в качестве затравки, поли(А)-полимераза последовательно присоединяет к нему остатки AMP из ATP по тому же самому механизму, что и другие ДНК- и РНК-полимеразы. Для эффективного функционирования поли(A)-полимераза требует наличия фактора CPSF, а также поли(A)-связывающего белка PAB II (poly(A)-binding protein II), который связывает полиаденилирующий комплекс с РНК после присоединения к ней, по крайней мере, десяти остатков А. В присутствии этих двух факторов поли(А)-полимераза сразу синтезирует поли(А)-последовательность полной длины по процессивному механизму. Гипотетическая структура элонгирующего комплекса представлена на рис. I.14.
Процессивное (непрерывное) полиаденилирование 3’-концов РНК происходит со скоростью ~ 25 нуклеотидов/с до тех пор, пока длина поли(А)-последовательности не достигнет ~ 250 нуклеотидов. После этого процессивная реакция прекращается, и происходит медленное дистрибутивное присоединение остатков AMP разными молекулами поли(А)-полимеразы. Предполагают, что элонгирующий белковый комплекс узнает длину синтезированной поли(А)-последовательности при участии фактора PAB II (см. рис. I.14). По этому механизму связывание определенного числа молекул PAB II с поли(А) прекращает элонгацию поли(А)-последовательности. Такой строгий контроль за длиной поли(А) на 3’-концах процессированных мРНК имеет большое значение для действия механизма, контролирующего время полужизни мРНК в цитоплазме. Без тщательного контроля над этим процессом с помощью селективного деаденилирования невозможно регулировать внутриклеточную деградацию мРНК, а вместе с тем и уровень экспрессии соответствующих генов с участием данного механизма.
Полиаденилирование является универсальным феноменом, играющим важную роль в процессинге и функционировании мРНК как прокариотических, так и эукариотических организмов. Однако сравнение механизмов полиаденилирования у этих групп организмов выявляет существенные различия, суммированные в табл. I.9.
Н. Саркаром (1997 г.) было высказано предположение о возникновении механизма полиаденилирования РНК. Он полагает, что поскольку у бактериальных и эукариотических ферментов, выполняющих аналогичные функции, не обнаружено гомологии в аминокислотных последовательностях, оба фермента возникли недавно из уже значительно дивергировавших функционально родственных предшественников. Такими предшественниками могли быть прокариотические и эукариотические тРНК-нуклеотидилтрансферазы, осуществляющие посттранскрипционный синтез последовательности CCA на 3’-концах тРНК, что по своему механизму близко к полиаденилированию. Подтверждением этого является обнаруженная значительная гомология между тРНК-нуклеотидилтрансферазой и основной поли(А)-полимеразой E. coli, а также между тРНК-нуклеотидилтрансферазой бактерии Sulfolobus shibatae, обитающей вгорячих серных источниках, и поли(А)-полимеразами эукариот. Следы такой эволюционной связи обнаруживаются и в современных митохондриях, где в результате полиаденилирования мРНК могут создаваться терминирующие кодоны. По мнению Саркара, различия между системами полиаденилирования прокариот и эукариот, представленные в табл. I.9, можно рассматривать в качестве продукта эволюционной дивергенции сходных биосинтетических функций в процессе независимого возникновения нового регуляторного механизма, обеспечивающего физиологические нужды уже глубоко различающихся групп организмов.
Таблица I.9