Мембраны возбудимых клеток имеют ионные каналы, которые могут находиться в двух состояниях: закрытом и открытом. Когда ионный канал открыт, то ионы свободно диффундируют через мембрану по градиенту их концентрации. Проницаемость мембраны для иона, диффундирующего через определенный тип канала, взаимосвязана с величиной мембранного потенциала. Процесс закрытия или открытия ионного канала обусловлен потенциалзависимыми воротными эффектами, которые возникают в ионном канале очень быстро и сохраняются кратковременно (1—2 мс). К такому типу потенциалзависимых ионных каналов относятся калиевые ионные каналы. Более медленные потенциалзависимые воротные эффекты имеют место в ионных каналах, которые могут находиться в трех состояниях: открытом, инактивированном и закрытом. Механизм инактивирова- ния ионного канала принципиально отличается от такового, который имеет место при открытии или закрытии (например, калиевого ионного кана
ла), тем, что канал вначале открывается, а затем инактивируется, и остается в таком состоянии некоторое время. К такому типу потенциалзависимых ионных каналов относятся натриевые ионные каналы мембраны возбудимых клеток, обусловливающие генерацию потенциала действия.
Потенциал действия нервной и мышечной клетки является основным проявлением в них процесса возбуждения. В состоянии покоя потенциалзависимые ионные каналы мембраны клеток возбудимых тканей закрыты, но сама мембрана находится в поляризованном состоянии, т. е. имеет л*ел*- бранный потенциал.
2.3.1. Мембранный потенциал покоя
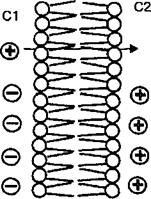
Мембранный потенциал покоя, или потенциал покоя, является результатом разности потенциалов между внутренней и наружной сторонами мембраны клетки (Ет). В покое внутренняя сторона мембраны клетки заряжена отрицательно по отношению к наружной, т. е. клеточная мембрана поляризована. Поляризация мембраны клеток заключается в следующем (рис. 2.8). Если мембрана клетки проницаема только для одного типа ионов, то в случае, когда концентрация этого типа ионов по обе стороны мембраны различна, они способны путем диффузии переходить через мембрану из среды с большей концентрацией (С7) в среду с меньшей концентрацией ионов (С2). В случае, если ионы заряжены положительно (см. рис. 2.8) как, например, ионы калия, то на стороне С2 мембраны накапливаются только положительно заряженные ионы. Положительный заряд ионов на стороне С2 мембраны будет препятствовать дальнейшему проникновению одноименных ионов, так как возникшее электрическое поле уравновешивает движущую силу концентрационного градиента положительных ионов на мембране. Электрические силы положительных ионов на стороне С2 мембраны будут притягивать отрицательные ионы со стороны С1 мембраны, для которых она непроницаема. При достижении равновесия концентрационные потенциалы ионов по обе стороны мембраны будут равны по величине, но противоположны по знаку их электрическому потенциалу. В результате по обе стороны мембраны возникает разность потенциалов. Возникновение на мембране разности потенциалов (Ет) описывается уравнением Нернста, которое позволяет рассчитать равновесный электрический потенциал для любого иона:
Ет = Zr с2
|
Рис. 2.8. Механизм возникновения потенциала Нернста.
Положительные ионы диффундируют из области высокой концентрации (С 1) в область низкой концентрации (С2). В результате одна сторона мембраны заряжается положительно, что препятствует дальнейшей диффузии положительных ионов на сторону С2. Между сторонами С1 и С2 возникает электрическое поле, которое уравновешивает движущую силу концентрационного потенциала, и устанавливается динамическое равновесие. Со стороны С1 на мембране формируется отрицательныйй заряд, а со стороны С2 — положительный. В целом, на мембране возникает разность потенциалов.
ность ионов, F - 96 500 Кл/моль — постоянная Фарадея, R ~ 8,31 Дж/ мольбК — универсальная газовая постоянная, Т — абсолютная температура системы, zF — заряд одного моля ионов, переносимых через мембрану.
Рассмотренный на рис. 2.8 пример свидетельствует о том, что потенциал на мембране клетки обусловлен ионами в том случае, если она для них проницаема. В отношении возбудимых клеток потенциал на их мембране может быть обусловлен ионами Na+, К? и СГ. В случае, если бы потенциал на мембране определялся только ионами калия, то его величина, согласно уравнению Нернста, должна равняться —90 мВ, принимая во внимание значение температуры тела человека (57 QQ, валентность иона {z ~ — 7), постоянную Фарадея (F == 96 500 Кл/моль) и универсальную газовую постоянную (R = 8,31Дж/молъКУ где К* и К*н — наружная и внутренняя концентрация ионов калия вне и внутри нейрона. В нейронах теплокровных животных отношение [7f/] к [А^] составляет порядка У28.
 | |||
 | |||
В случае, если бы мембрана клеток была пассивно проницаема только для ионов Na+ или только для ионов С1", то на ней устанавливался бы потенциал, равный +60 мВ и —80 мВ соответственно. При этом для хлора величина потенциала Нернста близка к реальной величине потенциала покоя нейрона, т. е. примерно —80 мВ. Однако в норме потенциал покоя на мембране нейронов составляет порядка -60 мВ. Причина отклонения потенциала на мембране нейрона от потенциала, рассчитанного по уравнению Нернста для каждого из названных ионов, заключается в том, что через мембрану нейрона постоянно движутся встречные потоки ионов К+, Na+ и СГ, каждый из которых имеет разный коэффициент проницаемости.
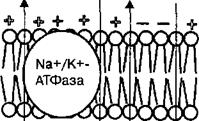
Потенциал покоя в возбудимых клетках. В возбудимых клетках между цитозолем и внеклеточной средой существует концентрационный градиент для ионов Na+ и К+, который поддерживается работой натрий-калиевого насоса, функционирующего за счет гидролиза энергии АТФ. Активность натрий-калиевого насоса создает разность потенциалов на мембране порядка —4 мВ, поэтому его называют электрогенным насосом. Основной функцией натрий-калиевого насоса является поддержание градиента концентрации ионов Na+ и К+ на мембране нейрона. В результате активности натрий-калиевого насоса концентрация ионов К+ с внутренней стороны мембраны клеток в 20—50 раз выше, чем с внешней стороны мембраны, а с внешней стороны мембраны в 8—10 раз выше концентрация ионов Na+. Поэтому ионы калия стремятся выйти из клетки, а ионы натрия — проникнуть внутрь. В покое мембрана возбудимых клеток проницаема в разной степени для нескольких ионов, т. е. ионов К+, Na+ и СГ. Это является причиной отклонения потенциала на мембране от потенциала, рассчитанного по формуле Нернста для каждого из этих ионов. В этом случае мембранный потенциал может быть описан уравнением Гольдмана—Ходжкина—Катца:
Ет = Alin ^[^1 +^[Ж+1 +/,с,[С4„~]
F Рк[ХЛ + PnANcC} + Pcz[ Cl„ ]'
где Em — мембранный потенциал; R — газовая постоянная; T — абсолютная температура; F— число Фарадея; Рк, PNn и Ра — константы проницаемости
мембраны для ионов К\ Na+ и СГ; /О /О /Ж7 /ВД /Ш и [С1вн]— концентрации ионов К+, Na+ и СГ снаружи (н) и внутри (вн) клетки.
Согласно формуле Гольдмана—Ходжкина—Катца концентрационный градиент каждого из ионов обусловливает величину мембранного потенциала. В покое величина мембранного потенциала пропорциональна проницаемости мембраны для ионов К+, Na+ и СГ, которая составляет — Рк: PNa: РС| = 1: 0,04: 0,45. Величина коэффициента проницаемости ионов калия свидетельствует о том, что концентрационный градиент положительно заряженных ионов калия направлен из клетки во внеклеточную среду. По своему характеру проницаемость ионов калия через мембрану нейрона носит диффузионный характер (ток утечки ионов калия) и потенциал покоя определяется, в основном, этим ионом. Во внутриклеточной среде возбудимых клеток содержатся различные анионы, которые не могут проникать через мембрану (белки, аминокислоты, органические кислоты, фосфаты, сульфаты). В результате избыток отрицательных ионов внутри клетки создает отрицательный заряд на поверхности мембраны с ее внутренней стороны. Выход ионов калия через мембрану из клетки создает избыток положительных ионов на внешней поверхности мембраны.
Концентрация ионов хлора по обе стороны мембраны возбудимых клеток находится в равновесном состоянии. Поэтому ионы хлора могут двигаться через мембрану в обоих направлениях с одинаковой скоростью. На этом основании ионы хлора могут не входить в уравнение Гольдмана- Ходжкина—Каца. Лишь при быстром нарушении равновесного состояния ионы хлора могут оказывать влияние на величину потенциала на мембране возбудимых клеток.
2.3.2. Потенциал действия возбудимых клеток
Потенциал действия возбудимых клеток представляет собой кратковременное фазное изменение знака потенциала на внутренней поверхности мембраны и проницаемости мембраны для ионов, вызванное внешним раздражением мембраны возбудимых клеток. Потенциал действия состоит из фаз деполяризации, реполяризации и следовых потенциалов.
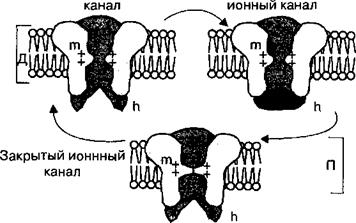
|
Рис. 2.9. Натриевый воротный ионный канал. При деполяризации мембраны канал находится в открытом или инактивированном состояниях (закрыты h-ворота). При поляризованной мембране, т. е. когда внутренняя поверхность мембраны имеет отрицательный заряд относительно наружной, ионный канал находится в закрытом состоянии (m-ворота закрыты).
Д — деполяризация мембраны, П — поляризация мембраны в состоянии покоя.
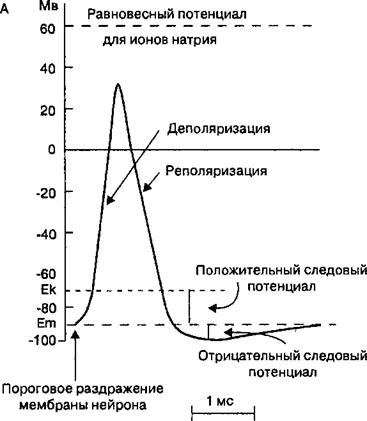
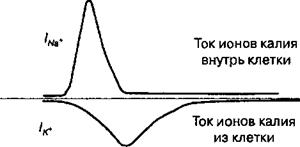 Рис. 2.10. Потенциал действия.
Рис. 2.10. Потенциал действия.
Изменения мембранного потенциала (А), которые называются потенциалом действия нервов и мышц, являются результатом изменения ионной проницаемости мембраны для Na’ и К* (Б). В зависимости от иона, участвующего в генерации потенциала действия, изменение ионной проницаемости мембраны происходит в результате открытия, закрытия или инакги- вирования ионных каналов. Большинство потенциалзависимых натриевых ионных каналов открывается, когда деполяризация достигает порогового уровня (Ek). После спонтанной инактивации этих каналов открываются калиевые ионные каналы, что вызывает реполяризацию мембраны. В период, когда натриевые ионные каналы находятся в инактивированном состоянии, мембрана клетки является рефрактерной. Натриевые ионные каналы функционируют примерно в 10 раз быстрее, чем калиевые.
ничего». Согласно этому закону величина потенциала действия остается постоянной, независимо от силы вызвавшего его стимула. Стимулом для генерации потенциала действия явля
 |
ется быстрая кратковременная деполяризация мембраны возбудимой клетки, которая называется критическим уровнемдеполяризащи (£к). Величина потенциала на мембране (Ет), необходимая для достижения критического уровня деполяризации, называется пороговым потенциалом (Ет—Ек), или порогом генерации потенциала действия (рис. 2.10). Амплитуда критического уровня деполя- ризациии мембраны нервного волокна и мышечной клетки в среднем меньше величины потенциала покоя на 15—20 мВ. Пороговый сдвиг мемциала происходит медленно в течение нескольких миллисекунд, то инак- тивационные ворота в натриевых ионных каналах инактивируются раньше, чем произойдет открывание достаточного количества активационных ворот. В результате натриевый ток (1„а) будет недостаточным для открытия потенциалзависимых натриевых каналов. Подобное явление называется аккомодацией мембраны В этом случае не будет возникать потенциал действия. Деполяризация мембраны начинается с активации инактивированных потенциалзависимых натриевых ионных каналов, и только после этого натриевые каналы открываются. Ионы ла + через открытые потенциал
зависимые натриевые каналы устремляются по концентрационному градиенту внутрь клетки (натриевый ток /№), заряжая внутреннюю поверхность мембраны положительным зарядом. При этом отношение коэффициентов проницаемостей мембраны для ионов калия и натрия составляет иную величину относительно состояния покоя: Рк: PNa = 1: 20, а генерация потенциала действия обусловлена высокой проницаемостью мембраны для ионов Na\ Деполяризация мембраны клетки увеличивает число открытых потенциалзависимых натриевых каналов, что существенно повышает натриевую проницаемость. В результате потенциал на мембране возбудимой клетки за 1—2 мс стремится достичь величины натриевого равновесного потенциала. При этом потенциал на внутренней поверхности мембраны меняет знак и его величина достигает +40 мВ (явление реверсии заряда мембраны).
Реполяризация. Во время реверсии заряда мембраны величина потенциала Нернста для ионов натрия приближается к их равновесному потенциалу, что вызывает через 1—2 мс после начала деполяризации инактивацию потенциалзависимых натриевых ионных каналов. При этом инактивацион-
ные ворота натриевых ионных каналов не открываются до тех пор, пока мембранный потенциал покоя не восстанавливается до своего исходного уровня. В результате прекращается входящий натриевый ток. Одновременно под влиянием деполяризации мембраны открываются потенциалзависимые калиевые ионные каналы и увеличивается выходящий калиевый ток, который стремится сместить мембранный потенциал в сторону калиевого равновесного потенциала и, таким образом, реполяризует мембрану (рис. 2.11). Мембранный потенциал приближается к величине потенциала покоя, но в мембране еще остаются открытыми ка
 | |||
 | |||
| |||||
| |||||
|
 |
|
 |
ваться ее нормальная возбудимость. Величина потенциала покоя мембраны нейронов составляет —60 мВ, а мембраны нервного волокна и скелетной мышечной клетки —90мВ, что свидетельствует о более высокой возбудимости сомы нейронов, чем их аксонов и, тем более, скелетных мышечных волокон.
При этом амплитуда потенциала действия в нервных волокнах составляет 110 мВ, что на +20 мВ превышает величину потенциала покоя. В скелетных мышечных клетках во время фазы деполяризации величина потенциала на мембране достигает максимального значения +30—40 мВ. Поэтому амплитуда потенциала действия в клетках скелетных мышц составляет 120—130 мВ. Эти различия в амплитуде реверсии заряда мембраны нервных и мышечных волокон обусловливают большую скорость протекания возбуждения в нервных волокнах, а следовательно, более высокую их лабильность по сравнению с мышечными волокнами.
2.2.3. Рефрактерный период в возбудимых клетках
В фазу деполяризации потенциала действия потенциалзависимые натриевые ионные каналы кратковременно открываются, но затем инактивируются h-ворота (см. рис. 2.9). В период инактивации натриевых ионных каналов возбудимые клетки не способны реагировать повышением натриевой проницаемости на повторный стимул. Поэтому во время фазы деполяризации мембрана не может генерировать потенциал действия в ответ на действие пороговых или сверхпороговых раздражителей. Это состояние называется абсолютной рефрактерностью, время которой составляет в нервных волокнах 0,5—1,0 мс, а в скелетных мышечных клетках —в среднем 2 мс. Период абсолютной рефрактерное™ заканчивается после того, как снижается количество инактивированных натриевых каналов и постепенно увеличивается количество натриевых каналов, находящихся в закрытом состоянии. Происходят эти процессы во время фазы реполяризации, когда уменьшению количества потенциалзависимых натриевых ионных каналов, находящихся в состоянии инактивации, соответствует период относительной рефрактерности. Период относительной рефрактерное™ характеризуется тем, что лишь некоторая часть потенциалзависимых натриевых ионных каналов переходит в закрытое состояние, а в силу этого порог возбудимости мембраны клетки имеет более высокие значения, чем в исходном состоянии. Поэтому возбудимые клетки в период относительной рефрактерности могут генерировать потенциалы действия, но при воздействии на них раздражителей сверхпороговой силы. Однако из-за небольшого количества потенциалзависимых натриевых ионных каналов, находящихся в закрытом состоянии, амплитуда генерируемых при этом потенциалов действия будет меньше, чем в условиях исходной возбудимости нервной или мышечной клетки.
В клетках возбудимых тканей максимальное количество генерируемых потенциалов действия в единицу времени обусловлено двумя факторами: длительностью потенциала действия и длительностью периода абсолютной рефрактерности после каждого импульса. На этом основании в физиологии формулируется современное понятие лабильности: чем меньше период абсолютной рефрактерности при возбуждении возбудимой ткани, тем выше ее функциональная подвижность или лабильность, тем больше в ней генерируется потенциалов действия в единицу времени.
При непрерывной стимуляции нерва электрическим током лабильность нерва зависит от частоты и силы раздражения. В зависимости от частоты и
силы раздражения нерва сокращение иннервируемой им мышцы может быть максимальной или минимальной амплитуды. Эти явления были названы соответственно оптимумом и пессимумом (Н. Е. Введенский). Максимальное (оптимально большое) сокращение мышцы возникает в том случае, если каждый последующий электрический стимул действует на нерв в периоде его состояния супернормальной возбудимости после предыдущего потенциала действия. Минимальное (или пессимальное) сокращение мышцы возникает в том случае, если каждый последующий электрический стимул действует на нерв, находящийся в периоде относительной рефрактерности после предыдущего потенциала действия. Поэтому значения оптимальной частоты раздражения нерва всегда меньше, чем значения пессимальной частоты раздражения.
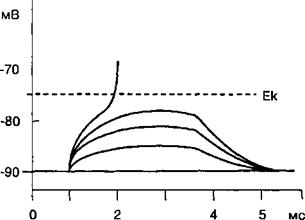
2.3.4. Локальный ответ мембраны возбудимых клеток
При действии электрического стимула подпороговой силы уменьшение мембранного потенциала оказывается недостаточным для того, чтобы натриевый ток (INa) стал больше, чем сумма калиевого выходящего тока или тока утечки ионов калия. При этом мембранный потенциал уменьшается
на несколько милливольт, но в результате реполяризации восстанавливается до величины мембранного потенциала покоя. Подобные пассивные или электротонические изменения потенциала на мембране, которые не достигают критического уровня деполяризации под действием деполяризующего электрического тока подпороговой величины, называются локальным ответом. Для локального ответа характерны следующие физиологические проявления на мембране нервной
 | |||
 | |||
|
 |
ствия кратковременного импульса электрического тока подвергаются обратному самостоятельному развитию. При действии на возбудимую мембрану серией подпороговых электрических стимулов происходит суммирование амплитуды локального ответа. Если амплитуда локального ответа достигает порога возбудимости мембраны, то потенциалзависимые натриевые ионные каналы открываются по закону «все или ничего» и происходит генерация потенциала действия. На рис. 2.12 показано суммирование амплитуды локального ответа по типу деполяризации при действии на мембрану нервного волокна серией подпороговых электрических стимулов. Локальный ответ нервного волокна всегда сопровождается повышением возбудимости мембраны клеток.