Передача возбуждения с отростка одной нервной клетки на отросток или тело другой нервной клетки возможна двумя способами: электрическим и химическим. Электрический способ передачи возбуждения осуществляется с помощью щелевых контактов (нексусов) в том случае, если мембраны клеток отделены щелью размером примерно 2 нм. Щелевые контакты распространены, например, в миокарде и гладких мышцах пищеварительного тракта и представляют собой межклеточные тубулярные белковые гидрофильные каналы, состоящие их шести молекул интегральных белков- коннексонов. Передача возбуждения электрическим способом осуществляется аналогично его проведению по нервным волокнам с помощью местных токов, возникающих между деполяризованным участком мембраны нервного волокна и поляризованным участком мембраны нервной клетки. Локальные токи деполяризуют мембрану нейрона до критического уровня, после чего возникает спонтанный процесс регенеративной деполяризации. Электрическая передача возбуждения осуществляется с высокой скоростью, близкой к скорости проведения возбуждения по нервным волокнам.
Основным способом передачи возбуждения между нейронами, а также между нервными и мышечными клетками является химический, который реализуется с помощью синапсов. Синапс — это специализированное по структуре и функции место контакта мембран между двумя возбудимыми клетками, которое обеспечивает передачу импульсов от одной возбудимой
клетки к другой с помощью молекул химических веществ, или медиаторов. Проведение возбуждения между нервом и мышцей осуществляется через нервно-мышечный синапс, который представляет собой место контакта окончания аксона нейрона с мембраной мышечного волокна. Функцией такого соединения является передача сигналов возбуждения от «-мотонейронов спинного мозга к скелетным мышечным волокнам, вызывающих сокращение мышцы.
2.5.1. Проведение возбуждения через нервно-мышечный синапс
Нервно-мышечный синапс образован мембраной терминали аксона, или пресинаптического окончания мотонейрона спинного мозга, и мембраной мышечного волокна, или постсинаптической мембраной, между которыми имеется синаптическая щель (рис. 2.15). Постсинаптическая мембрана нервно-мышечного синапса называется концевой пластинкой, поскольку представляет собой специализированную часть мембраны мышечного волокна, имеющую складчатый вид, что существенно увеличивает площадь мембраны, контактирующей с аксолеммой нервной терминали. Кроме того, постсинаптическая мембрана содержит N-холинорецепторы и фермент ацетилхолинестеразу, которая разрушает медиатор ацетилхолин. Исходя из структурной организации нервно-мышечного синапса, передачу возбуждения с нерва на мышцу можно разделить на два физиологических механизма: пресинаптический и постсинаптический. Функция нервно-мышечного синапса заключена в передаче очень небольших по амплитуде потенциалов действия, достигающих двигательных нервных окончаний, на мышечные волокна и инициации, таким образом, сокращения во всей скелетной мышце.
Синаптические Пресинаптическая Терминаль аксона
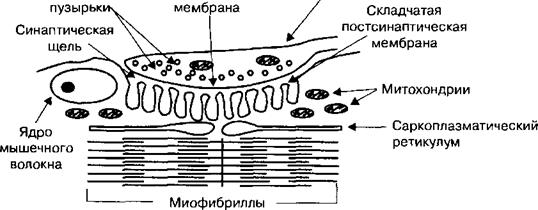
Рис. 2.15. Схема нервно-мышечного синапса.
Функцией соединения является трансформация энергии нервного импульса в кванты ацетилхолина, который выделяется из терминали в синаптическую щель с последующим превращением химического сигнала в электрический на мембране мышечного волокна. На складчатой постсинаптической мембране находятся ионотропные холинорецепторы, которые, связываясь с ацетилхолином, открывают натриевые ионные каналы постсинаптической мембраны. Деполяризация постсинаптической мембраны электротонически открывает потенциалзависимые натриевые ионные каналы сарколеммы и, таким образом, в ней генерируются распространяющиеся потенциалы действия.
25. L1. Пресинаптический механизм
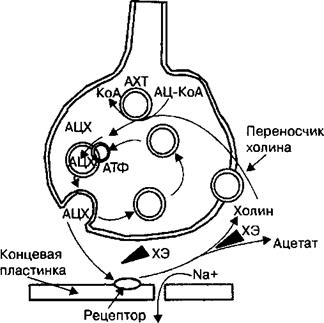
|
 В синаптических пузырьках терминали аксона, иннервирующего мышцу, содержится медиатор ацетилхолин. Ацетилхолин синтезируется непосредственно в нервном окончании из холина и ацетил-Ко-А при участии фермента ацетилтрансферазы, Наряду с медиатором в везикулах содержатся различные ионы, АТФ, ферменты и другие вещества, участвующие в выделении медиатора в синаптическую щель. В каждом синаптическом пузырьке содержится примерно 10~4 молекул ацетилхолина. Количество ацетилхолина, высвобождаемое из одной везикулы, принято называть квантом медиатора. Без ресинтеза ацетилхолина в двигательном нервном окончании запаса его молекул в синаптических пузырьках достаточно для проведения примерно 10 000 импульсов. Везикулы пресинаптической терминали находятся в активной и депонированной формах. Активные везикулы непосредственно участвуют в передаче возбуждения через нервно-мышечное соединение, т. е. в выделении медиатора в синаптическую щель. Молекулы медиатора депонированной формы везикул постоянно пополняют
В синаптических пузырьках терминали аксона, иннервирующего мышцу, содержится медиатор ацетилхолин. Ацетилхолин синтезируется непосредственно в нервном окончании из холина и ацетил-Ко-А при участии фермента ацетилтрансферазы, Наряду с медиатором в везикулах содержатся различные ионы, АТФ, ферменты и другие вещества, участвующие в выделении медиатора в синаптическую щель. В каждом синаптическом пузырьке содержится примерно 10~4 молекул ацетилхолина. Количество ацетилхолина, высвобождаемое из одной везикулы, принято называть квантом медиатора. Без ресинтеза ацетилхолина в двигательном нервном окончании запаса его молекул в синаптических пузырьках достаточно для проведения примерно 10 000 импульсов. Везикулы пресинаптической терминали находятся в активной и депонированной формах. Активные везикулы непосредственно участвуют в передаче возбуждения через нервно-мышечное соединение, т. е. в выделении медиатора в синаптическую щель. Молекулы медиатора депонированной формы везикул постоянно пополняют
|
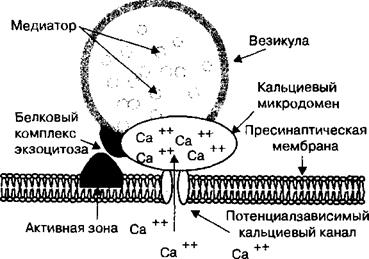
 После того как нервный импульс достигает пресинаптического окончания в фазу деполяризации потенциала действия, в мембране окончания открываются потенциалзависимые кальциевые ионные каналы, и ионы Са2+ пассивно по градиенту концентрации поступают внутрь нервного окончания из внеклеточной среды. Движение ионов Са2+ обусловлено тем, что в покое их концентрация в цитоплазме нервного окончания составляет порядка 10“7 М, а на внешней поверхности его мембраны, благодаря активности Са2+-АТФазы, поддерживается более высокая концентрация ионов кальция, чем внутри. В момент открытия кальциевых ионных каналов в активных зонах пресинапти- ческой мембраны формируются микродомены (рис. 2.17). Микродомен представляет собой область высокой концентрации ионов Са2+ вблизи устья кальциевого ионного канала. В момент появления микродомена белки мембраны везикулы способны связывать везикулярную мембрану с активной зоной пресинаптического окончания. При этом образуется специфический белковый комплекс, состоящий из везикулы и элементов активной зоны пресинаптической мембраны, которые связаны между собой специализированными белками экзоцитоза. Этот комплекс называется сек- ретосомой. Формирование секретосомы является основным процессом, обеспечивающим экзоцитоз ацетилхолина в нервном окончании.
После того как нервный импульс достигает пресинаптического окончания в фазу деполяризации потенциала действия, в мембране окончания открываются потенциалзависимые кальциевые ионные каналы, и ионы Са2+ пассивно по градиенту концентрации поступают внутрь нервного окончания из внеклеточной среды. Движение ионов Са2+ обусловлено тем, что в покое их концентрация в цитоплазме нервного окончания составляет порядка 10“7 М, а на внешней поверхности его мембраны, благодаря активности Са2+-АТФазы, поддерживается более высокая концентрация ионов кальция, чем внутри. В момент открытия кальциевых ионных каналов в активных зонах пресинапти- ческой мембраны формируются микродомены (рис. 2.17). Микродомен представляет собой область высокой концентрации ионов Са2+ вблизи устья кальциевого ионного канала. В момент появления микродомена белки мембраны везикулы способны связывать везикулярную мембрану с активной зоной пресинаптического окончания. При этом образуется специфический белковый комплекс, состоящий из везикулы и элементов активной зоны пресинаптической мембраны, которые связаны между собой специализированными белками экзоцитоза. Этот комплекс называется сек- ретосомой. Формирование секретосомы является основным процессом, обеспечивающим экзоцитоз ацетилхолина в нервном окончании.
Белки экзоцитоза локализованы как на мембране везикул (синапсин, синаптотагмин, синаптобревин), так и на пресинаптической мембране (синтаксин, синапсоассоциированный белок). Белок синаптотагмин является кальциевым рецептором. После связывания ионов Са2+ с синаптотаг- мином и последующего взаимодействия синаптобревина с синаптосомас- социированным белком и синтаксином пресинаптической мембраны происходит связывание везикулы с активной зоной пресинаптической мембраны. Экзоцитоз медиатора обеспечивается за счет АТФазной активности специфического цитозольного белка. При высвобождении ацетилхолина в синаптическую щель мембрана везикулы полностью сливается с пресинаптической мембраной и медиатор выходит в синаптическую щель через так называемую пору. Белки экзоцитоза в синапсах периферической нервной системы являются мишенями для токсинов при их действии на организм человека и животных. Например, ботулиновый, столбнячный и другие токсины способны нарушать процессы формирования секретосомы, что обусловливает появление специфического симптомокомплекса при отравлении.
2.5.1.2. Диффузия ацетилхолина через синаптическую щель нервно-мышечного синапса
Синаптическая щель нервно-мышечного синапса заполнена аморфной сетью соединительной ткани, в которой находится фермент ацетилхолин- эстераза. Молекулы этого фермента синтезируются в мышечной клетке и фиксируются к ее сарколемме со стороны синаптической щели.
Ацетилхолин путем экзоцитоза поступает в синаптическую щель и затем диффундирует к постсинаптической мембране в течение 1 мс. Во время диффузии ацетилхолина через синаптическую щель примерно 73 от выделившегося количества медиатора разрушается ферментом ацетилхолин- эстеразой на ацетат и холин, а 2/3 молекул ацетилхолина достигает постсинаптической мембраны и связывается с N-холинорецепторами. Ацетилхолин после отделения от рецептора расщепляется ацетилхолинестеразой на холин и ацетат. Синтез, хранение и высвобождение ацетилхолина в нервно-мышечном соединении показаны на рис. 2.16. Активность ацетилхоли- нэстеразы столь высока, что период полураспада молекул ацетилхолина не превышает 1 мс. Инактивация ацетилхолина в синаптической щели исключает многократное связывание одних и тех же молекул медиатора с хо- линорецепторами постсинаптической мембраны, что предотвращает возможность непрерывного сокращения мышц или так называемой мышечной тетании.
2.5.1.3. Постсинаптический механизм
Постсинаптический механизм передачи возбуждения через нервно-мышечный синапс представляет собой трансформацию количества выделившегося ацетилхолина в число потенциалов действия, которые генерирует сарколемма мышечного волокна. В постсинаптической мембране нервно-мышечного соединения расположены ионотропные N-холинорецепторы (чувствительны к никотину). Плотность этих рецепторов выше, чем в постсинаптической мембране любого синапса нервной системы (1 • 104 на 1 мкм2). Молекулы ацетилхолина кратковременно (не более 2 мс) взаимодействуют с N-холинорецепторами, в результате в постсинаптической мембране открываются натриевые и калиевые ионные каналы. Ионы Na+ по электрохимическому градиенту перемещаются из синаптической щели внутрь мышечного волокна, а ионы К+ — в обратном направлении. Трансмембранный ток для этих ионов рассчитываются по закону Ома:
4 = §к(Ет - £к); = gNa(^ “
где 1К и INa — трансмембранный ток ионов калия и натрия, gK и gNa — проницаемость постсинаптической мембраны соответственно для ионов калия и натрия, Ет — потенциал покоя постсинаптической мембраны, Ек и ENa — равновесный калиевый и натриевый потенциалы.
Поскольку gNa = gK, а электродвижущая сила (Em — ENa) для натрия больше (равновесный потенциал для натрия +60 мВ, а для калия — 90 мВ), входящий ток ионов натрия преобладает над выходящим калиевым током, в результате происходит деполяризация постсинаптической мембраны нервно-мышечного синапса, которая называется потенциалом концевой пластинки. Продолжительность потенциала очень мала — не более 1—2 мс, а его амплитуда варьирует в зависимости от количества высвобождающегося ацетилхолина. При этом между амплитудой потенциала концевой пластинки и количеством высвобождающегося ацетилхолина имеется прямая зависимость.
Потенциал концевой пластинки электротонически распространяется вдоль мышечного волокна. При этом, если потенциал концевой пластинки достигает по величине порога возбуждения мышечной мембраны (—15— 20 мВ), то он вызывает в ней генерацию распространяющихся потенциалов действия. В результате возбуждение с нервного окончания передается на мышечную клетку. Если пресинаптического окончания достигает один потенциал действия, то в синаптическую щель нервно-мышечного синапса высвобождается в среднем 100 квантов ацетилхолина. В этом случае возникает потенциал концевой пластинки, величина которого достаточна для того, чтобы вызвать генерацию потенциалов действия в мышечной мембране, прилегающей к концевой пластинке. Даже при снижении амплитуды потенциала концевой пластинки на 75 % он способен вызывать возбуждение в мышечном волокне, что является фактором высокой надежности процесса передачи возбуждения в нервно-мышечном синапсе. Надежность функции нервно-мышечного синапса обусловлена также значительным количеством высвобождаемого из нервного окончания ацетилхолина и высокой плотностью N-холинорецепторов на концевой пластинке.
Как только потенциалы действия, распространяясь по мембране мышечного волокна, достигают t-системы, а ионы Са2+ высвобождаются из саркоплазматического ретикулума, в мышце начинается сокращение.
2.5.1.4. Восстановительные процессы структуры мембраны и функции нервно-мышечного синапса после передачи возбуждения
После завершения синаптической передачи в нервно-мышечном синапсе ионы Са2+ с помощью Са2+-АТФ-азы первично активным транспортом выводятся из цитоплазмы нервного окончания во внеклеточную среду. Процесс восстановления исходного градиента концентрации ионов Са2+ между аксоплазмой нервной терминали и внеклеточной средой осуществляется с помощью вторично активного транспорта при участии белка Na+/Ca2+ обменника. Молекулярная структура пресинаптической мембраны восстанавливается в результате эндоцитоза везикул, участвовавших в синаптической передаче возбуждения. В нервном окончании образовавшиеся везикулы заполняются вновь синтезированными молекулами ацетилхолина. На постсинаптической мембране, благодаря работе Ма+/К2+-АТФазы, восстанавливаются ионные градиенты Na+ и К+, в результате возбудимость постсинаптической мембраны возвращается к исходному уровню.
2.5.2. Проведение возбуждения через аксосоматический синапс
В нервной системе проведение возбуждения осуществляется между отростками нейронов и между отростками и телами нервных клеток. Так, проведение возбуждения с аксона одного нейрона на аксон другого происходит при участии аксо-аксональных синапсов. Передача импульсов от аксона нейрона на на дендриты другой нервной клетки осуществляется аксоденд- ритными синапсами, а от одних дендритов на другие —с помощью дендро-дендритных синапсов. К наиболее часто встречающемуся в нервной
Микротрубочки
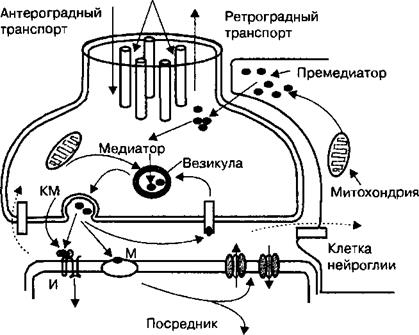
Рис. 2.18. Схема строения и функции химического синапса.
Предшественник медиатора попадает в терминаль нейрона из крови или ликвора, под действием специализированного фермента превращается в медиатор и активным транспортом накапливается в синаптических везикулах. Медиатор после экзоцитоза или после выделения ко- медиатора (КМ) взаимодействует с ионотропным (И) или метаботропным (М) рецептором постсинаптической мембраны. В результате изменяется проницаемость этой мембраны для ионов Na+, К+, Са2+, СГ и возникает локальная де- или гиперполяризация путем непосредственного изменения ионной проницаемости, либо при участии вторичного посредника. Медиатор разрушается ферментом в синаптической щели, захватывается активным транспортом в пресинаптическое окончание либо в клетки нейроглии (пунктирные стрелки).
системе пути передачи импульсов между нейронами относится аксосома- тический синапс, т. е. от аксона одного нейрона на тело другого. Количество синапсов на дендритах и теле одного нейрона достигает нескольких тысяч. В качестве примера может быть рассмотрен аксосоматический синапс, т. е. синапс между окончаниями одной нервной клетки и телом другой (рис. 2.18).
Аксосоматический синапс образован пресинаптическим окончанием аксона, синаптической щелью, за которой расположена постсинаптическая мембрана. Передача импульсов в аксосоматических синапсах осуществляется с помощью медиаторов, которые называются нейротрансмиттерами, В химической передаче возбуждения между нейронами участвуют различные типы нейротрансмиттеров: глутамат, ацетилхолин, дофамин, норадреналин, адреналин, серотонин, гамма-аминомасляная кислота (ГАМК), нейропептиды. При передаче возбуждения через аксосоматический синапс в постсинаптическом нейроне возникает два типа ответных реакций: быстрые и медленные. Быстрые реакции в постсинаптических нейронах возникают в случаях, если медиатор связывается с рецепторами ионных каналов постсинаптической мембраны и, непосредственно вызывая их открытие, в миллисекундный интервал передает возбуждение на постсинаптический нейрон. При этом возникают быстрые реакции постсинаптической мембраны в виде увеличения ионной проницаемости и ее деполяризации. К быстрым реакциям увеличения ионной проницаемости постсинаптической мембраны аксосоматических синапсов относится открытие ионных каналов постсинаптической мембраны после образования внутриклеточных вторичных посредников (описаны ниже). При этом типе быстрой реакции нейротрансмиттеры (адреналин, норадреналин, дофамин, гистамин, серотонин) связываются на постсинаптической мембране с рецепторами системы G-белков. В результате активации системы G-белков в постсинаптическом нейроне образуются внутриклеточные вторичные посредники, которые через рецепторы ионных каналов открывают их и увеличивают таким образом ионную проницаемость постсинаптической мембраны аксосоматических синапсов. При передаче возбуждения через аксосоматический синапс возникают медленные или нейромодуляторные реакции постсинаптических нейронов. Они также обусловлены взаимодействием нейротрансмиттеров с рецепторами системы G-белков постсинаптической мембраны, но в этом случае при участии внутриклеточных вторичных посредников возникают медленные, а именно — метаболические реакции постсинаптических нейронов. Данный способ передачи информации через синапсы нервной системы присущ большой группе нейротрансмиттеров, которые называются нейропептидами.
По аналогии с нервно-мышечным синапсом в аксосоматическом синапсе импульс предается с помощью пре- и постсинаптического механизмов.
2.5.2.1. Функция пресинаптического окончания нейронов
В нервных терминалях локализовано два типа везикул — мелкие и крупные. Мелкие везикулы терминалей нейронов центральной и периферической нервной системы содержат разные группы медиаторов: амины (ацетилхолин, дофамин, норадреналин, серотонин и гистамин), аминокислоты (глютамат, глицин, гамма-аминомасляная кислота) и пурины (АТФ, аденозин). Мелкие везикулы в пресинаптической терминали находятся в активной и депонированной формах. Активные везикулы непосредственно участвуют в экзоцитозе медиатора, депонированная форма везикул пополняет пул активной по мере ее использования в синаптической передаче (рис. 2.19). Медиаторы накапливаются в везикулах с помощью механизма активного транспорта. Крупные или пептидные везикулы нервных терминалей центральной и периферической нервной систем содержат нейроактивные пептиды (субстанция Р, вазопрессин, окситоцин, кортиколиберин, тиреолиберин, соматолиберин, гонадолиберин, энкефалины, ангиотензин- 2 и др.). В отличие от мелких везикул пептидные синтезируются в соме нейронов, наполняются молекулами пептидов и транспортируются с помощью аксотока в нервное окончание. Функции медиаторов мелких и крупных везикул взаимосвязаны между собой в процессе синаптической передачи: первые обеспечивают химическую передачу возбуждения через синапс, а вторые — модулируют, т. е. усиливают или уменьшают этот процесс. Генерация потенциала действия на мембране пресинаптической терминали активирует кальцийзависимый процесс освобождения или экзоцитоза медиатора. Быструю, кратковременную (доли миллисекунды) и локальную передачу возбуждения между нейронами обеспечивает экзоцитоз классических нейротрансмиттеров, содержащихся в мелких везикулах нервной терминали. Экзоцитоз нейропептидов из нервных терминалей может осуществляться в течение многих секунд и минут при активации различных внутриклеточных энзимных систем. Благодаря этой особенности экзоцитоза нейропептидов, они могут модулировать экзоцитоз класси-
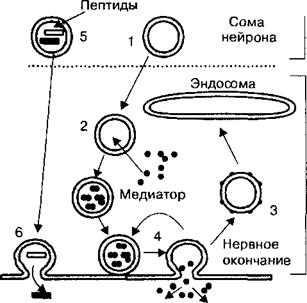 Рис. 2.19. Везикулярный механизм экзоцитоза медиатора из нервной терминали.
Рис. 2.19. Везикулярный механизм экзоцитоза медиатора из нервной терминали.
Везикулы (1) формируются в теле нейрона, транспортируются в нервное окончание, где заполняются медиатором (2). При экзоци- тозе, с помощью механизма полного слияния и с последующим образованием покрытых везикул (3), запасы медиатора пополняются через стадию эндосом. При неполном слиянии и повторном заполнении медиатором везикулы сразу могут участвовать в эк- зоцитозе (4). Крупные везикулы заполняются пептидами в теле нейрона (5), их эк- зоцитоз происходит в других зонах нервного окончания (6).
ческих нейротрансмиттеров в ней
ронах на протяжении продолжительного интервала времени, усиливая или замедляя передачу импульсов через аксосоматический синапс. В аксосоматическом синапсе экзоцитоз классических медиаторов осуществляется в активных зонах пресинаптического окончания. Функциями активных зон в нервных терминалях (как и в нервно-мышечном синапсе) является мобилизация медиатора из депонированной формы в активную при участии элементов цитоскелета. В пределах активной зоны пресинап- тической мембраны происходит взаимодействие между везикулами и местом экзоцитоза медиаторов. При участии активных зон пресинаптической мембраны экзоцитоз происходит только в момент повышения концентрации ионов кальция в цитоплазме пресинаптической терминали. Наконец, активные зоны нервной терминали участвуют в процессах восстановления целостности синаптических пузырьков, заполнения их молекулами медиаторов, что обусловливает число везикул, находящихся в активной и депонированной формах.
2.5.2.2. Пресинаптический механизм проведения возбуждения
Пресинаптический механизм проведения возбуждения осуществляет трансформацию энергии нервных импульсов в кванты выделяющегося медиатора. В фазу деполяризации потенциала действия в мембране нервной терминали открываются потенциалзависимые кальциевые ионные каналы, и ионы кальция по концентрационному градиенту поступают из внеклеточной среды внутрь нервного окончания. Ионы Са2+ вблизи внутренней поверхности пресинаптической мембраны образуют микродомен (см. рис. 2.17) и связываются со специфическим везикулярным белком кальциевым сенсором — синаптотагмином. Последовательное взаимодействие специфических белков экзоцитоза, локализованых на мембране везикул (синапсин, синаптотагмин, синаптобревин) и на пресинаптической мембране (синтаксин, синапсоассоциированный белок) связывает везикулу с активной зоной пресинаптической мембраны, формируя секретосому. При этом мембрана везикулы полностью сливается с пресинаптической мембраной, и медиатор освобождается в синаптическую щель через пору. В синапсах нервной системы экзоцитоз медиаторов может проходит через постоянную или временную пору. В первом случае происходят полное слияние мембран везикулы и пресинаптического окончания и высвобождение всего количества молекул медиатора из везикул. Во втором случае не происходит полного слияния мембран везикулы и пресинаптической терминали. При образовании временной поры освобождается только часть медиатора в синаптическую щель, а затем везикула отделяется от активной зоны и возвращается в цитоплазму нервной терминали. Этот механизм позволяет одной и той же везикуле без пополнения запаса медиатора неоднократно участвовать в экзоцитозе.
После высвобождения молекулы нейротрансмиттера диффундируют от мест экзоцитоза к постсинаптической мембране. Во время диффузии часть медиатора может подвергаться ферментному разрушению в синаптической щели или захватываться обратно внутрь нервного окончания. Постсинаптической мембраны могут достигать все или только часть молекул нейротрансмиттеров, что обусловливает соответственно большее или меньшее количество передаваемых через аксосоматический синапс импульсов.
2.5.2.3. Пресинаптическая регуляция экзоцитоза медиаторов
Пресинаптическая регуляция экзоцитоза медиаторов осуществляется с помощью специфического типа ионного канала, который идентичен калиевым ионным каналам мембран возбудимых клеток, но открываются ионами кальция. Такие ионные каналы пресинаптической мембраны называются калъцийзависимыми калиевыми каналами. Эти ионные каналы расположены в активных зонах пресинаптической мембраны нервной терминали в непосредственной близости от кальциевых ионных каналов. Физиологические свойства подобного типа ионных каналов заключается в том, что они быстро активируются при увеличении концентрации ионов кальция на внутренней поверхности пресинаптической мембраны, т. е. в момент формирования микродомена. При открытии кальцийзависимых калиевых ионных каналов происходит выход из нервного окончания ионов калия по концентрационному градиенту. Это локально реполяризует пресинаптиче- скую мембрану нервной терминали, что ограничивает длительность фазы деполяризации в тех участках пресинаптической мембраны, где происходит высвобождение медиатора. В результате уменьшения длительности фазы деполяризации пресинаптической мембраны нервной терминали расположенные в ней кальциевые ионные каналы открываются на меньший отрезок времени. Это ведет к уменьшению количества ионов кальция, поступающих внутрь нервной терминали, а следовательно, уменьшаются количество высвобождаемого экзоцитозом медиатора и передача сигналов через синапс.
Освобождение медиаторов из нервных терминалей изменяется (модулируется) субстанциями, содержащимися в интерстициальной жидкости, окружающей терминаль. Механизм регуляции экзоцитоза медиаторов из нервных терминалей осуществляется через рецепторы, локализованные на пресинаптической мембране. Эти рецепторы (N-холинергические, глюта- матные, ГАМК-ергические и пуринергические Р2-рецепторы) ассоциированы с ионными каналами и могут активироваться медиатором, который выделяется из этого же нервного окончания, либо медиаторами, высвобождающимися из близлежащих нервных окончаний.
Вещества, которые оказывают модуляторный эффект на передачу импульсов в синапсах нервной системы, в результате которой увеличивается или уменьшается частота потенциалов действия постсинаптических нейронах, называются нейромодуляторами.
Экзоцитоз ацетилхолина из нервной терминали стимулируется связыванием выделившегося в этом же синапсе ацетилхолина с N-холинорецепто- рами пресинаптической мембраны аксосоматического синапса. Глютамат- ные рецепторы пресинаптической мембраны аксосоматического синапса (NMDA, агонист N-метил-О-аспартат; АМРА, агонист альфа-амино-3- гидрокси-5-метил-4-изоксазолпропионовая кислота и каинатные — агонист каиновая кислота) участвуют в разных синапсах в стимуляции или торможении экзоцитоза других медиаторов. Например, NMDA-рецепторы пресинаптической мембраны нервных терминалей, оканчивающихся на нейронах коры головного мозга, стимулируют процесс экзоцитоза медиатора ацетилхолина, а в нервных терминалях гиппокампа — медиатора норадреналина.
На пресинаптической мембране нервных терминалей нейронов центральной нервной системы распространен А! тип аденозиновых рецепторов, которые связываются с нейромодулятором аденозином. Концентрация аденозина во внеклеточной среде возрастает по мере активации постсинаптического нейрона. Функцией типа Ai аденозиновых рецепторов пресинаптической мембраны нервной терминали является уменьшение экзоцитоза медиаторов (ацетилхолина, дофамина), что снижает активность постсинаптических нейронов, из которых в окружащую их внеклеточную жидкость поступает аденозин. Тип А2А-аденозиновых рецепторов локализован на пресинаптической мембране нейронов тех отделов мозга, которые регулируют движения человека при участии медиатора дофамина (хвостатое ядро и подушка таламуса). Активация А2а рецепторов пресинаптической мембраны стимулирует высвобождение ацетилхолина из нервных терминалей и вызывает усиление функции постсинаптического нейрона.
25.2.4. Постсинаптический механизм проведения возбуждения
Постсинаптический механизм проведения возбуждения в аксосоматиче- ском синапсе нервной системы осуществляет передачу нервного импульса на постсинаптический нейрон {быстрые реакции). Передача нервного импульса от одного нейрона к другому осуществляется в течение короткого отрезка времени (мс). Этот процесс на этапе постсинаптического механизма передачи возбуждения осуществляется при участии ионотропных рецепторов постсинаптической мембраны аксосоматического синапса. Ионотропные рецепторы представляют собой часть молекулярной структуры хе- мозависимых ионных каналов (натриевых, калиевых, кальциевых) постсинаптической мембраны нейронов. Количество ионотропных рецепторов на постсинаптической мембране нейронов является регулируемой величиной: оно повышается в ответ на длительное воздействие на мембрану нейрона малой концентрации молекул медиаторов (явление сенситизации). Напротив, число рецепторов на постсинаптической мембране понижается в ответ на длительное воздействие на постсинаптическую мембрану медиаторов в высокой концентрации (явление десенситизации). Вещества, которые связываются с рецепторами хемозависимых ионных каналов постсинаптической мембраны, называются лигандами. Лиганды подразделяются на агонисты и антагонисты. Первые активируют, а вторые блокируют активность рецепторов хемозависимых ионных каналов постсинаптической мембраны.
Результатом взаимодействия медиаторов с ионотропными рецепторами постсинаптической мембраны является ее кратковременная деполяризация или гиперполяризация постсинаптической мембраны. Деполяризация постсинаптической мембраны в области одного синапса называется возбуждающим постсинаптическим потенциалом (ВПСП). Химические синапсы, в которых при передаче возбуждения возникают ВПСП, называются возбуждающими. По аналогии медиатор, вызывающий возникновение ВПСП, называется возбуждающим медиатором. Возникновение ВПСП обусловлено открытием преимущественно натриевых ионных каналов. При ВПСП происходит деполяризация постсинаптической мембраны и уменьшение порога ее возбудимости. Амплитуда ВПСП прямо зависит от количества молекул медиатора взаимодействующих с рецепторами постсинаптической мембраны, а также последовательностью выделения квантов медиатора из пресинаптического окончания.
Результатом взаимодействия медиаторов с ионотропными рецепторами хемозависимых ионных каналов постсинаптической мембраны может быть ее кратковременная гиперполяризация. В результате понижается возбудимость постсинаптического нейрона и в нем не происходит генерация потенциала действия. Гиперполяризация постсинаптической мембраны в области одного синапса называется тормозным постсинаптическим потенциалом (ТПСП), такие химические синапсы называются тормозными. По аналогии медиаторы, вызывающие ТПСП, называются тормозными медиаторами. Взаимодействие тормозного медиатора с рецепторами постсинаптической мембраны открывает хлорные и калиевые ионные каналы, что резко повышает ионную проводимость постсинаптической мембраны для ионов хлора (/с/) или калия (7^). В соответствии с электрохимическими градиентами ионов калия и хлора по обе стороны от постсинаптической мембраны, ТПСП представляет собой сдвиг потенциала на постсинаптической мембране в сторону увеличения его величины. В результате происходит гиперполяризация постсинаптической мембраны и увеличение порога ее возбудимости.
При передаче возбуждения через аксосоматические синапсы на мембране одного нейрона одновременно возникают ВПСП и ТПСП. При этом происходит сложение (суммация) амплитуды ВПСП и ТПСП многих синапсов. Если в условиях суммации ВПСП и ТПСП суммарная амплитуда ВПСП достигает критического уровня деполяризации, то в мембране аксонного холмика нейрона электротонически открываются потенциалзависимые натриевые ионные каналы и генерируется потенциал действия. В результате возбуждение передается от нервных терминалей пресинаптического нейрона на постсинаптический нейрон.
При передаче возбуждения через аксосоматический синапс с пресинаптического нейрона на постсинаптический происходит синаптическая задержка скорости проведения электрического импульса. Синаптическая задержка в возбуждающих аксосоматических синапсах представляет собой время от момента прихода потенциала действия к пресинаптическому окончанию до начала деполяризации постсинаптической мембраны, т. е. ВПСП. Время синаптической задержки в возбуждающих синапсах в среднем составляет 0,5 мс, и оно прямо зависит от скорости процессов экзоцитоза медиатора из нервного окончания, диффузии медиатора через синаптическую щель к постсинаптической мембране, соединения медиатора с рецепторами постсинаптической мембраны и последующего увеличения ионной проницаемости постсинаптической мембраны. Время синаптической задержки обусловливает функциональную подвижность или лабильность химического синапса. Лабильность химического синапса ниже, чем нервного волокна, но выше, чем мышечного.
2.5.25. Функции метаботропных рецепторов постсинаптической мембраны аксосоматического синапса
На постсинаптической мембране аксосоматических синапсов нервной системы локализованы хемозависимые метаботропные рецепторы. Функция хемозависимых метаботропных рецепторов постсинаптической мембраны состоит в регуляции метаболической активности постсинаптических нейронов при передаче возбуждения через химический синапс (медленная реакция). Эта функция метаботропных рецепторов постсинаптической мембраны обусловлена свойствами нескольких высокомолекулярных семейств G-белков (Gs, Gi и Gq) либо низкомолекулярных семейств G-белков (ras, rho, rab и др.), с которыми ассоциированы метаботропные рецепторы постсинаптической мембраны (рис. 2.20).
Как показано на рис. 2.20, метаботропный рецептор постсинаптической мембраны находится на стороне молекулы G-белка, обращенной в синаптическую щель. Соединение нейротрансмиттера с хемозависимым метаботропным рецептором постсинаптической мембраны вызывает активацию G-белков. При этом тример G-белков, состоящий из а-субъединицы, ассоциированной с Ру-димером, диссоциирует на а-субъединицу и Ру-димер с последующим образованием комп