ВВЕДЕНИЕ

Первые генетические манипуляции, применяемые к Toxoplasma были выполнены с помощью химического мутагенеза. Эти исследования были впервые в 1970-е годы Elmer Pfefferkorn (Pfefferkorn и Pfefferkorn, 1976; Пфефферкорн, 1988), который усовершенствовал протоколы воспроизводимо культивировать тахизоиты в системе культуры ткани и мутагенизации, выбирать, и, наконец, клонировать паразитов путем лимитирующего разведения. На основании этих протоколов, ряд химически индуцированных мутантов были использованы, чтобы наметить нуклеотидные пути биосинтеза пара-сайтов. Эти исследования имеют решающее значение для создания протоколов для генетических скрещиваний у кошки (Pfefferkorn и Pfefferkorn, 1980). Крестики могут быть использованы для отображения данного фенотипа в однократный или многократный геноме локусы. Это классический вперед генетический подход играет важную роль для сопоставления факторов вирулентности и
анализировать структуру населения Toxoplasma и эволюции. (См главы 3 и 14 для дальнейшего обсуждения этих вопросов.)
Обратной генетики подход, интро-Duces чужеродной ДНК в паразитов, была достигнута с помощью электропорации. Изначально переходный транс-fection плазмидной ДНК, содержащие репортерные гены, фланкированных T.gondii, 5' и 3'фланкирующие последовательности позволили экспрессию репортерных генов, используемых для характеристики элементов, контролирующих транскрипцию. Эта методика была использована быстро идентифицировать и проверить несколько селективных маркерные гены, которые затем открыли путь для стабильной трансформации и развитий бесценного арсенала средств, связанных с трансфекцией ДНК. Широкий спектр положительных и отрицательных селективных маркеры был приспособлен для homolo-рекомбинации Гуся, приводящей к аллельным заменам и генных нокаутам. Кроме того, не гомологичны
Токсоплазма. Модельные Apicomplexan-перспективы и методы, под редакцией Weiss & Kim
ISBN-13: 978-0-12-369542-0
ISBN-10: 0-12-369542-2
Copyright © 2007 Elsevier Ltd.
Все права воспроизведения в любой форме зарезервированы.

| ГЕНЕТИЧЕСКАЯ МАНИПУЛИРОВАНИЕ токсоплазма | |||
| Генетическая крест | Micronemes | ||
| векторы экспрессии | Внутренняя мембрана комплекс | ||
| Rhoptries | |||
| Эписомный вектор | Apicoplast | ||
| Плотная гранула | |||
| Джин нокаутов | паразитофорной вакуоль | ||
| инсерционный мутагенез | Сценический конкретный ЭЕТ | ||
| РНК антисмысловый | геномная | ||
| Tet-индуцируемый система | Microarrays | ||
| комплементация клонирование | протеомики |


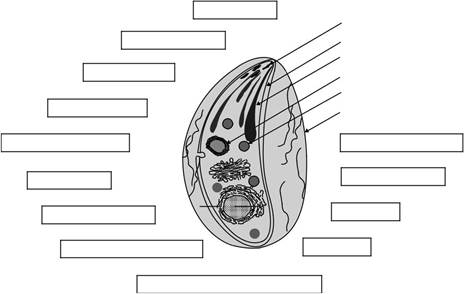
Подпись меченых мутагенеза
Рисунок 15.1 Источники информации и манипулированию стратегий. Схематическое изображение внутриклеточного паразита с субклеточными структурами и органеллами, а также перечнем инструментов, в настоящее время для функционального анализа. Рисунок из модифицированного Soldati, Д. и Мейснера, M. (2004). Токсоплазма модельного организма для Apicomplexans. В: Маларии Паразиты: геномов и молекулярной биологии. Норвич: Horizon Press, стр 135-167..
векторы случайных интеграции были разработаны, чтобы выразить трансгенов и в качестве стратегии случайного инсерционного мутагенеза.
Недавнее завершение проекта генома секвенирования Toxoplasma (WWW. Toxodb.org) и наличие других apicomplexan геномов для сравнения доставка беспрецедентного количества захватывающей информации. В этой новой области постгеномную, доступность Т. гондий на несколько генетических манипуляций подходов и исследований с высокой пропускной способностью делает его очень привлекательным и мощная система, чтобы улучшить наше понимание фундаментальной биологии APICOM-plexan паразитов. Рисунок 15.1 обобщает имеющиеся источники информации и экспериментальных подходов. Там нет ограничений на Опознавательная-катиона соответствующих генов, а также частичное или полное отсутствие барьера экспериментально разгадке их биологическую функцию на относительно больших масштабах.
Цель этой главы состоит в том, чтобы резюмировать и описать стратегии, связанные с трансфекции ДНК, в том числе самых последних приобретений, а также предоставить список наиболее полезных протоколов, реагентов и штаммов, доступных для исследователей.
механика получения трансгенных Паразитов
Переходная трансфекция
Успешная манипуляция Toxoplasma генома в решающей степени зависит от эффективности трансфекции ДНК. Электропорация была и до сих пор остается метод выбора для введения ДНК в тахизоиты. Важно отметить, что сочетание этого метода со средствами массовой информации имитируя цитозольное
| Механика ТРАНСГЕННЫХ паразитов ИЗГОТОВЛЕНИЯ |
ионный состав клеток (cytomix) дает лучший коэффициент выживаемости (ван ден Хофф и др., 1992). Протокол, которая первоначально была создана с использованием BTX электро-porator, привело к эффективности временной экспрессии, которые колебались от 30 до 50 процентов (Soldati и Бутройда, 1993). Оптимальные параметры, выбранные на BTX электропораторе были установлены для штамма RH (типа I, вирулентный штамм) и были немного изменены для цист-образующих штаммов (ME49 и Prugniaud; штаммы II типа). Было отмечено, что часто киста образующие штаммы менее поддаются генетической манипуляции, вероятно, связано с несколькими факторами.
Для того, чтобы контролировать эффективность трансфекции, chlorampheni-ацетил-трансферазы Col (CAT) и βгалактозидазы первоначально были использованы в качестве репортерных генов, и subse-тельно βлактамазы, щелочной фосфатазы и Светик люциферазы (LUC). Эти ферменты классифицирует-чески используется в качестве репортеров, потому что их деятельность может контролироваться с большой чувствительностью и количественным способом. Кроме того, эти ферменты отсутствуют в эукариотических клетках, что приводит к практически без фоновой активности.
Что интересно, βлактамазы и щелочной фос-phatase проявляют никакой активности в пределах паразита, вероятно, связано с наличием ингибиторов, и может быть использован для изучения секреторного пути и количественно секрецию паразита (Карстен и др., 1998;. Chaturvedi и др, 1999), LacZ-активность может быть тез-Ured с использованием колориметрического анализа, который преобразует желтый ХЛОРФЕНОЛ красно-β-D-галактопиранозида (CPRG) субстрат в красный продукт с использованием спектрофотометра оптическую плотность при 570 нм (Seeber и Бутройда, 1996). Этот колориметрический анализ считывания можно наблюдать в живых паразитов с использованием культуральной среды без фенола красного и в мульти-луночных планшетах, позволяющих (на высоком уровне пропускной способности) скрининг эффективности препарата против паразита (McFadden и др., 1997).
Правоверных экспрессии гена-репортера требует адекватной 5' и 3'фланкирующие последовательности, которые являются производными от генов гондий T.. Эти фланкирующие последовательности должны содержать элементы управления, необходимые для привода оптимального уровня transcrip-ции. Monocistronic природа транскрипции в T.gondii, способствовали выявлению промоторных элементов, которые, как правило, в непосредственной близости к
начало сайта транскрипции. Многочисленные векторы, пригодные для трансфекции в настоящее время доступны, и, так как они демонстрируют различный диапазон прочности промотора и стадия специфичности, они могут быть выбраны-ately Ассигнования на в соответствии с целью эксперимента.
Было отмечено, что часто прочность, и, вероятно, также время экспрессии по отношению к клеточному циклу, критически повлиять на результатах эксперимента, особенно при исследовании-ным внутриклеточной локализации данного продукта гена. Например, избыточная экспрессия microneme белков часто приводит к накоплению в раннем отделении секреторного пути-пути, или неплотности в паразитофорную вакуоль (Сольдать и др., 2001). Использование природного промотора настоятельно рекомендуется, если результаты трансгенного гена Expres-SION токсичности или mistargeting (Gubbels и др., 2006b).
Конститутивный уровень экспрессии может быть получен с использованием векторов, полученные из TUB1 (α-tubulin), DHFR (дигидрофолатредуктазы), ROP1 (rhoptry белок, 1), MIC2 (microneme белок 2), несколько ГРЫ(плотные гранулы белок) и HXGPRT (гипоксантин-гуанин-phosphori bosyl трансферазы) гены. Прочность этих и других промоторов, не очень систематически сравнивать, но промоторы ГРА и MIC2 являются самыми сильными, TUB1 и ROP1 промоторы являются промежуточными, в то время как DHFR-TS является слабым промотором.
Выражение Стадия конкретных может быть достигнуто с использованием 5-фланкирующих последовательностей генов стадии конкретных, и до сих пор ни на одном этапе конкретных регуляторные элементы не были отображены в 3'UTR последовательности. Выражение тахизоит конкретным присваиваются векторами, полученных из SAG1 (поверхностного антигена 1), ENO1 (енолаза 1), и LDH1 (лактат дегидро-genase 1) генов. В противоположность этому, векторы, сконструированные из BAG1 (брадизоитный антиген 1), ENO2 (енолаза 2) или генов SAG4 придают экспрессию в стадии брадизоитный исключительно. Детальный промотор анализ и идентификация цис-действующие элементы были проведены только для ограниченного числа генов (Soldati и Бутройда, 1995; Mercier и соавт, 1996;. Bohne и соавт, 1997;. Ян и Parmley, 1997; Matrajt и др., 2004; Kibe и др, 2005).. Глава 16 предусматривает обсуждение регуляции экспрессии генов.
394 ГЕНЕТИЧЕСКАЯ МАНИПУЛИРОВАНИЕ токсоплазма
В дополнение к элементам промотора, последовательность Характеристика осуществляется на мРНК, также вносят вклад в успех трансфекции. Информация о последовательности получена из 5' и 3'нетранслируемые области, вероятно, влияет на экспрессию генов, но этот уровень регуляции не был тщательно расследоваться на сегодняшний день. 3'UTR является важным элементом, как тран-scription падает до менее чем 10 процентов, когда такой элемент не входит. В Plasmodium, частичное удаление из 3' UTR регионы были использованы для модуляции уровня экспрессии основных генов, предлагая способ анализа их функции (Thathy и др., 2002).
В инициирующем кодоне, консенсусная последовательность называется "последовательность Козака распознается рибосомами как благоприятные последовательности для инициирования перевода. Подборка обильных выраженных генов в T.gondii, была использована для установления последовательности инициации консенсуса трансляционной gNCAAaATGg, которая подобна, но не идентична последовательности Козака, найденной в высшем эукариоте (Seeber, 1997). Несколько генов, включая GFP были изначально очень трудно выразить с помощью своей нативной последовательности, но отсутствие экспрессии была решена путем генерации слитых на N-конце (Striepen и др., 1998). Эти наблюдения указывают на
a значительное влияние последовательностей N-концевой амино-кислот в экспрессии рекомбинантного белка. Систематический анализ, направленный на оценку важности аминокислоты после инициации метионина подтвердил существование N-концевого правила в Т. гондий (Matrajt и др., 2002a). Аминокислоты, такие как Ala, Glu и Asp придают высокий уровень экспрессии трансгена.