При формальном подходе к проблеме иннервации половой сферы можно, например, констатировать, что центростремительная дуга эрекционного рефлекса начинается от эрогенной зоны на головке полового члена что эрекционный центр расположен в крестцовых сегментах спинного мозга (в отличие от эякуляторного, локализующегося в поясничных) и что центробежное колено эрекционного рефлекса посредством связано с гладкой мускулатурой, регулирующей просвет артерий кавернозных тел полового члена. Если же к этой проблеме подойти операционально, т. е. с целью разобраться в функциональном взаимодействии различных анатомических образований нервной системы, обеспечивающих всю последовательность физиологических проявлений полового акта, то на этом пути сексолог очень скоро встретит огромные трудности. Приведенное выше описание спинального центра эрекции к связанных с ним афферентной и эфферентной дуг дает крайн& одностороннее и потому искаженное представление об истинной иннервации эрекции. Как известно, эрекция в большинстве случаев возникает под влиянием импульсов, формирующихся на церебральном уровне, в результате дистантных воздействий
|
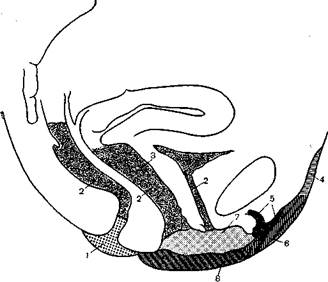
Рис. 28. Схема чувствительной иннервации половой сферы женщины [342].
1 — гатиз паетоггЪсийаНз т{еп9г пегу! рийепй! соттишз; 2 — пегуиз Ьаетогг-поМаЦз тесИаИз; 3 — пегуие уа81паНз; 4 — пегуиз Шо-теитаНз, пегуиз 1Ко-Ьуро-ВазШоиз; 5 — пегуиз йогваНя'вШт-ми- к — »о™,.~ 1„г,1_1--пегуиз уа^аз; 4 — пегуиз Шо-теиШа118, пегтид Шо-Ьуро-5 — пегуиз йогеаШ сЩогШз; 6 — гатиз ГаЫаНз ап1егтг пепа зрегта-Ыс1 ехгегп!; 7 — пегуиз уе8Ши1из уа^тае еЬ иге1Ьгае (пегуиз 1аЫаЦз роз4егюг); 8 — гатиз реппеиз цепа рийепсН соттишз.
(зрительных, обонятельных и слуховых), а подчас только посредством образов памяти, и эти воздействия способны вызвать
эрекцию даже при полном отключении афферентной дуги спи-
нального рефлекса (так же как в других случаях полный
поперечный перерыв спинного мозга в грудном отделе не препятствует возникновению эрекции, провоцируемой с соответствующих сегментарных Зон [125]). Более того, Мюллер [465] наблюдал эрекции у 2 больных с утратой всех видов чувствительности на половых органах, обусловленной полным разрушением крестцового отдела спинного мозга. Он же, разрушив в эксперименте нижние сегменты спинного мозга у собак-самцов,
вызывал у них затем эрекцию в присутствии находящихся в течке самок.
Аналогичный феномен получен Гольтцем [369]. Представленное Мюллером объяснение этих фактов может быть принято лишь для части случаев; оно сводится к тому, что эрекционный центр локализуется вне спинного мозга и что эрекция, как и другие вазомоторные реакции, регулируется экстрасшшально, от симпатического ствола (рис. 30). Постав
ленное ограничение в свою очередь выходит за рамки физиоло
|
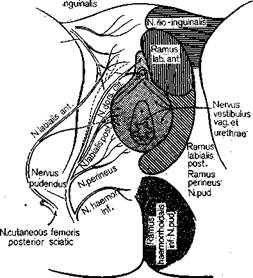
Рис. 29. Фронтальная схема распределения чувствительных нервов промежности женщины (справа) и схема ветвления магистральных нервных стволов (слева) [342].
Таким образом, анализ сложных отношений, свойственных половой сфере, показывает, что даже представленное на картах Дикинсона (см. рис. 28, 29) распределение периферической иннервации при бесспорной его специфичности, будучи включено в функциональный контекст, перекрывается явлениями более высокого порядка, что, в частности, отражается в наблюдениях над индивидуальными особенностями эрогенных зон [449, 450, 105, 213, 71]. При взаимодействии центра и периферии оба эти полюса оказывают друг на друга влияние, порождающее подчас весьма своеобразные отношения. Так, Мастере и Джонсон наблюдали нескольких женщин, у которых изолированное тактильное раздражение сосочков молочных желез (являвшихся у них ведущей эрогенной зоной) без каких бы то ни было генитальных воздействий вызывало наступление оргазма раньше, чем половой акт, проводимый без раздражения ареол сосков. Нами наблюдалась женщина, у которой, вне зависимости от длительности фрикций, оргазм не наступал до тех пор, пока мужчина не вводил ей палец в анальное отверстие.
Приведенные факты делают неприменимым в клинической сексопатологии традиционное представление о нервном центре как об анатомически узко локализованном скоплении ганглиоз-ных элементов, которое характеризуется абсолютной стабильностью пространственных и функциональных параметров.

Рис. 30. Схема иннервации мужских половых органов [342].
, а — чувствительные нервы; б — двигательные нервы; в — симпатические преганглионарные нервные волокна; г— постганглионарные нервные волокна 1—поперечное сечение спинного мозга в верхне-поясничном отделе- 2 — поперечное сечение нижне-крестцового отдела спинного мозга; 3 — нижний брыжеечный узел; 4 —аортальное сплетение (сплетение брюшной аорты); 5 — подчревное
• сплетение; 6 — семенные пузырьки; 7 — сплетение семявыводящего протока- 8 — мочевой пузырь; 9 —простата; 10 - симфиз; И —N йогзаНз решя- 12 — яичко 13 —т. ЪиШо-сауегпозиз; 14 — п. регопаеиз; 15 — сфинктер уретры.
Поэтому излагаемая здесь сексологическая концепция базируется не на анатомо-локализационистских постулатах, а на тех динамических представлениях о взаимодействии центра и периферии, которые разработаны школой П. К. Анохина, начиная сего публикации 1935 г. [6]: «Говоря о „центрах", в первую очередь подразумевают те нервные образования, которые являются в полном смысле этого слова мозговыми концами различных периферических нервных стволов. Эти нервные стволы являются непосредственной связью любого периферического органа, воспринимающего или эффекторного, с ганглиозными элементами, которые могут лежать в различных отделах центральной.нервной системы....Но описанные центры, являясь входными и выходными воротами для разнообразных циркулирующих по центральной нервной системе импульсов, связаны между собой 'через целый ряд промежуточных нервных образований, в которых циркуляция нервного импульса, благодаря их многосторонним связям, представляется особенно комплексной. Эти промежуточные нервные образования, к которым относятся преимущественно подкорковые образования и кора, с точки зрения "теории центров усложняют течение нервного процесса и нервные проявления организма только благодаря своему суммарному функционированию».При традиционном подходе свойства отдельных центров считаются строго анатомически фиксированлыми, и те ассоциационные волокна, которые осуществляют соединение между ними, в значительной степени отражают ту же специфичность: «Функциональная специфичность определенных нервных образований есть изначальное их свойство и ни при каких условиях не изменяется. Вот те основные положения, которых обязательно придерживается, сознательно или бессознательно, каждый клиницист и нейрофизиолог, стоящий на строго локализационной точке зрения» {6, стр. 65—66]. Понятие «центр» получило новое, функциональное определение в трудах П. К. Анохина и его сотрудников и в работах А. Б. Когана [123, 124] и других современных исследователей. По определению А. Б. Когана: «Нервный центр — это многоуровневая структура. В ее основании лежат наиболее простые, элементарные объединения нейронов, из которых строятся объединения второго порядка, которые в свою очередь объединяются в структуры третьего порядка и т. д.» [123]. П. Г. Костюк подчеркивает два основных принципиальных новоприобрете-ния, возникающих в результате интегрирования такого рода клеточных ансамблей: 1) объединение индивидуальных нейронов в системы, обладающие новыми свойствами, отсутствующими у входящих в их состав единиц предыдущего порядка; 2) то, что активность каждого составного элемента определяется не только влияниями, поступающими по непосредственным афферентным входам каждого элемента, но и состоянием других элементов системы [133].
Тот же примат функционально-динамических отношений между гибко приспосабливающимися к меняющимся требованиям клеточными объединениями над принципом абсолютно предопределенного диктата локализационной структурности формулируется и школой Н. П. Бехтеревой: «Исследования взаимодействия групп нейронов внутри одной нейронной популяции в первом приближении подтвердили представительность именно нейронного ансамбля, а не нейрона как звена системы обеспечения деятельности, и показали возможность развития различных отношений между нейронными ансамблями в процессе деятельности вплоть до противоположных состояний» (25].
Для сексопатолога, поскольку все виды рекреационных проявлений сексуальности суть поведенческие акты, в которых раскрываются высшие психические проявления личности, особую ценность в обсуждаемом круге закономерностей приобретает следующее суждение одного из создателей научной нейропсихологии А. Р. Лурия: «Современная психология подходит к высшим психическим процессам но как к изолированным „функциям" или более неразложимым „способностям", а как к сложным функциональным системам, которые достигают поставленной задачи целым комплексом способов или средств. Эти функциональные системы подвижны. Иначе говоря, система средств, с помощью которых может быть решена та или иная задача, может изменяться. Вначале работа функциональных систем носит развернутый характер (как это, например, имеет место у ребенка на ранних этапах обучения письму), затем она автоматизируется, свертывается» (156].
Физиология основных проявлений сексуальности мужчины
Эрекция (лат. егесИо, от еп^еге — поднимать) — увеличение полового члена в объеме по сравнению с состоянием покоя (приблизительно в три раза) и приобретение им механической твердости, необходимой для введения во влагалище и обеспечивающей проведение полового акта.
Эрекция наряду с эякуляцией — один из самых характерных феноменов мужской сексуальности, который в силу приданной ему символичности положен в основу фаллического культа (493, 524]. Один из основных парадоксов эрекции — в чрезвычайной ее чувствительности ко всякого рода функциональным психо-травмирующим воздействиям вопреки тому факту, что из всех основных проявлений мужской сексуальности именно эрекции отличаются наибольшей защищенностью от повреждающих факторов органической природы и наибольшей стабильностью (например, в онтогенезе эрекции выявляются задолго до либидо, эякуляции и оргазма).
Эрекция — рефлекторно-сосудистый акт; сосудистая природа ее была доказана более 200 лет назад голландским анатомоми физиологом Ренье де Граафом [370]. Применив свою методику инъекции контрастной массой сосудов трупа, он доказал, что в основе эрекции лежит наполнение пещеристых тел полового члена кровью, а не скопление в них воздуха, как полагал Гален. Некоторые авторы, в частности Чяшапп (1883), отождествляли вазомоторный механизм эрекции с венозным застоем, полагая, что в основе эрекции лежит затруднение оттока крови по отводящим венам полового члена, сдавливаемым луковично-пещеристой (и седалищно-пещеристыми мышцами; в то же время, благодаря сокращению, которые действуют наподобие рычага, сосудистый эффект усиливается чисто механическим приподниманием члена. Подобному допущению, однако, противоречили и длительность эрекции, и отсутствие неприятных ощущений, сопутствующих пассивному застою при накладывании жгута на конечность, и наблюдаемое при эрекции местное повышение температуры вместо понижения, отмечаемого при застое. Применив в эксперименте на собаках сфигмографию полового члена в сочетании с кимографической записью уровней внутрисосудистого давления на артериальном и венозном участках, Франсуа Франк доказал, что основной механизм эрекции составляет усиление артериального притока, а замедление оттока по венам играет лишь вспомогательную роль. Из анализируемых им кривых явствует, что давление возрастает сначала в артериях, и лишь после того — в венах [358]. И, наконец, Легро [433] перевязкой вен у корня полового члена показал, что затруднение венозного оттока может вызвать лишь некоторое набухание пещеристых тел, очень далекое от настоящей эрекции.
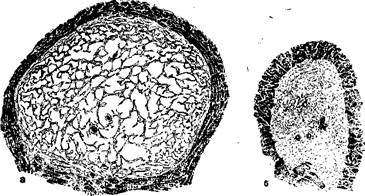
По описанию сосудистого механизма эрекции, оставленному Л. Я. Якобзоном [279]: «Кровь при эрекции действует не как сплошная масса жидкости, находящаяся под большим давлением, а распределена по многочисленным щелям пещеристых тел; причина этого заключается, по-видимому, в растягивающей и разрывающей силе, которая проявляется в оболочке всякого пузыря вследствие существующего внутри него давления... Этим обстоятельством можно, вероятно, объяснить то замечательное явление, что член может достигнуть твердости, наверное превышающей плотность аорты, хотя этой твердостью он обязан давлению крови, которое, однако, в аорте выше». И действительно, как доказал Левей [437], артериальное давление внутри пещеристых тел при эрекции достигает '/з—2/з давления в сонной артерии. Описанные Л. Я. Якобзоном отношения наглядно иллюстрируются на рис. 31. При этом выявляется еще один парадокс механизма эрекции, который заключается в том, что предельная ригидность, придающая кавернозным телам на высоте эрекции деревянистую плотность, достигается снятием мышечного спазма (см. рис. 27)
 .
.
]Рис. 31. Изменение объема пещеристых тел:
а — поперечный разрез через правое пещеристое тело полового члена, инъецированное фиксирующим раствором до состояния эрекции; б — аналогичный участок левого пещеристого тела в спавшемся состоянии. Та же степень увеличения [508].
В дополнение к сказанному необходимо упомянуть описанные Экснером [356] радиальные тяги, возникающие при эрекции, которые способствуют раскрытию как просвета уретры (рис. 32),
Ряд существенных различий в архитектонике пещеристых тел полового члена с одной стороны и пещеристого тела уретры — с другой (по относительному содержанию мышечных и эластических волокон, мощности белочной оболочки, расположению мышечных пучков и пещеристых лакун), заставили многих исследователей, в том числе Дикинсона [342] и А. И. Тюкова [248], провести разграничение между собственно пещеристыми телами полового члена и губчатым пара уретральным телом уретры.
Указанные различия проявляются в там, что кавернозное тело уретры, а следовательно, и головка полового члена - (см. рис. 22) даже на высоте эрекции оказываются менее напряженными, что создает более благоприятные условия для прохождения семени через уретру и придает головке полового члена роль буфера, смягчающего травматизацию женских гениталий: «Эректильная ткань головки члена всегда остается мягкой и податливой, хотя и увеличивается при эрекции в объеме; отмеченная эластичность головки облегчает ее проникновение во влагалище и предупреждает возможность повреждений. Она также предохраняет заостренные и ригидные концы кавернозных тел пениса как от получения разрушений (при случайном
|
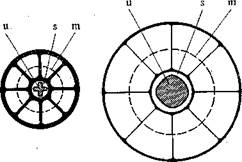
Рис. 32. Схема раскрытия просвета уретры под влиянием радиальных тяг, возникающих при заполнении кровью пещеристых пространств согриз сауег-позит игеШгае [356]:
а — согриз сатегпозит иге№-гае в спавшемся состоянии; б — то же в состоянии эрекции. II — уретра; 3 — слизистая оболочка уретры; т — окружность, проходящая на середине толщины согриз са-тегпозшп игеШгае.
столкновении с твердыми телами), так и от причинения травм при их посредстве» [342].
Кан, представивший указанные отношения на схеме (рис. 33), назвал механизм эрекции триумфом биомеханики, превзойти который не может надеяться ни один инженер: мягкие ткани превращаются в отличающееся предельной твердостью тело; это состояние поддерживается в течение длительного времени, не вызывая тягостных ощущений; приток крови уравновешивается соответствующим оттоком (при увеличении кровотока через пещеристые тела в 6—8 раз); шесть железистых образований взаимодействуют, выдерживая строго отрегулированный ритм и последовательность, и при этом в середине созданной повышенным давлением структуры остается незатрудненный проход для семенной жидкости. По определению А. И. Тюкова, эрекцию можно рассматривать как периодически возникающую физиологическую регионарную гипертонию [249].
Как отмечает Л. Я. Якобзон, переполнение члена кровью при эрекции распространяется и на перепончатую часть уретры, семенной бугорок и шейку мочевого пузыря. При этом отмечено, что сильная эрекция делает мочеиспускание невозможным или крайне его затрудняет. 'УУеЬег (1846) объяснял это набуханием семенного бугорка, который якобы заполняет весь просвет предстательной части уретры и, таким образом, замыкает ее. Однако, по справедливому замечанию Уокера [527], если бы это было так, то было бы непонятно, каким образом при полном закрытии мочеиспускательного канала в него проникают семя и секреты семенных пузырьков и предстательной железы. Далее Уокер показал [528], что семенной бугорок при налитии его сосудов не перекрывает просвет мочеиспускательного канала. В действительности, как это продемонстрировали рентгенологические исследования Цейсля и Хольцкнехта [536], мочеиспусканию при эрекции препятствует непроизвольное сокращение внутреннего сфинктера мочевого пузыря.

Рис. 33. Схема сосудистой архитектоники эрекции [409].
1—5 — сосудистый аппарат пещеристого (губчатого) тела уретры; а, Ь, с — сосудистый аппарат пещеристых тел полового члена.
Рефлекторная природа эрекции была доказана в 1839 г., получившим периферическим раздражением члена у животного с перерезанным спинным мозгом как эрекцию, так и эякуляцию. Затем Экхард [349] вызвал в эксперименте у собаки эрекцию прямым раздражением, а также продемонстрировал утрату возможности получения эрекции тактильным раздражением полового члена после перерезки. П. В. Никольский [469] подтвердив данные своих предшественников, отметил активное сокращение сосудов полового члена после перерезки пп. еп§еп1оз и наступление эрекции в результате раздражения их дистальных отрезков (скорость вытекания капель крови из перерезанных кавернозных тел при этом увеличивалась). Франк [358] показал, что сосудодвигательные нервы полового члена берут начало не только от крестцовых корешков, как это было описано по его данным, сосудодвигательный иннерваторный аппарат полового члена происходит из крестцового и поясничного отделов и составляется нервами, которые большой частью заканчиваются. По накопившимся к этому времени экспериментальным наблюдениям, возбуждение п. Франк показал, что оба были правы, так как нерв содержит оба сорта волокон и, в зависимости от способа раздражения, может быть и сосудосжимателем, и сосудорасширителем. В ситуациях, неблагоприятных для половых проявлений, а также по окончании полового акта, нервные импульсы, напротив, поступают на симпатический конечный путь и через подчревное сплетение воздействуют на вазоконстрикторы полового члена, подавляющие эрекцию. Таким образом, периферические ганглии, расположенные в пограничном симпатическом стволе, нижнем брыжеечном, подчревном и других сплетениях, являются «центрами» сосудистого тонуса, уровень активности которых в различных ситуациях то повышается, то понижается, подобно тому как изменяется влияние блуждающего нерва на деятельность сердечных узлов. Основываясь на своих экспериментальных данных, Гольтц пришел к выводу, что головной мозг представляет высшую инстанцию, интегрирующую посредством межцентральных нервных путей все «низшие механически-рефлекторные центры», выполняющие лишь «черную работу» сближения.
Эякуляция
Эякуляция (лат. е]аси1аНо— выбрасывание, извержение), или семяизвержение—выделение эякулята (продуктов деятельности семенников и придаточных половых желез).
Эякуляция, как и эрекция, акт рефлекторный, но гораздо более сложный, ибо относительно однозначному сосудистому компоненту эрекции здесь соответствует ряд компонентов двигательной и выделительной природы, развертывающихся в определенной последовательности, способствующей смешиванию различных секретов, что является необходимым условием для обеспечения жизнеспособности и активности спермиев. Лоде [435] и Экснер [356] установили, что у человека в серии эякуляций, следующих друг за другом с короткими перерывами, сперматозоиды исчезают в третьей или четвертой порции и вновь обнаруживаются в эякулятах, полученных после 2—3 суток полового покоя. Не удовлетворившись этими данными, поскольку получавшееся в экспериментах Лоде и Экснера опорожнение могло затрагивать лишь уровни семявыносящих протоков, а обнаруживавшиеся через трое суток сперматозоиды могли поступать в соответствующие отрезки половых путей не из семенников, а из придатков, Тутхилл и Янг [518], проследив в эксперименте на морских свинках за судьбой частиц туши, инъецированных в головку придатка, установили, что для продвижения спермиев из головки придатка в начальную часть семявыносящего протока требуется не менее 2—3 нед, и высказали предположение, что этому продвижению может способствовать активность ресничек выносящих канальцев. Наличие постоянного медленно протекающего продвижения сперматозоидов от головки к хвосту придатка, наблюдаемое даже при отсутствии копулятивной активности, способствует дозреванию спермиев {535].
Эякуляция как таковая начинается с активного мышечного сокращения семявыносящих протоков и разделяется на 2 фазы. Первая фаза — выведение семени, совершается непроизвольно под действием импульсов, следующих из сегментов по соединительным ветвям (гапп сопшшшсап^ез) симпатического ствола через нижний брыжеечный узел и подчревное сплетение [466, 496]; постганглио-нарные волокна распределяются по сомявьшосящим протокам на всем проикжсчши, начиная от придатков и включая ампулярную часть и частично семенные пузырьки, а также обеспечивают внутренний сфинктер мочевого пузыря [310] и констрикторы артерий полового члена [496], Фаза выведения знаменует переключение на качественно новый вид двигательной активности — секрет яичек, скопившийся до этого в результате пассивного и медленного проталкивания выделенных ранее порций яичкового секрета новыми, поступающими из придактов (под влиянием свойственной этим последним перистальтики), с началом первой фазы эякуляции в значительно возросшем темпе сокращениями мышечных стопок семявыводящих протоков активно перемещается в предстательную часть уретры. Поступление же эякулята в рагз ргозЪаИса игеЙиае, действуя как стимулятор парасимпатической и соматической иннервации, вызывает вторую фазу — фазу выбрасывания (изгнания, или извержения) уже не яичкового секрета, а эякулята. Вторая фаза эякуляции совершается еще более энергично, так как в ней к действию непроизвольных парасимпатических импульсов, следующих из сегментов 82 — 84, проходящих через внутренние срамные нервы, а также через интрамуральные сплетения некоторых тазовых органов (в частности, через и воздействующих на гладкую мускулатуру семенных пузырьков и предстательной железы, присоединяются мощные сокращения поперечно-полосатой мускулатуры, и вовлекающие всю как произвольную, так и непроизвольную мускулатуру тазового бедра. По данным Кафемана [408], действие аппарата эякуляции у отдельных лиц, достигает такой силы, что семенная жидкость извергается па расстояние более метра.
Первые наблюдения сокращений семяновыносящих протоков принадлежат Ксег (1851), который, производя гальванизацию их стенок на трупе, констатировал, что они «очень энергично укорачиваются и сокращаются». Важно подчеркнуть, что движения эти, как подтвердили и последующие исследования 1321, 316, 420, 84], не идентичны целенаправленной перистальтической волне, движущейся от одного конца трубки к другому. В частности, Бемингхаус, которому удалось заснять процесс эякуляции у морской свинки на кинопленку [316], показал, что содержимое член перемещается в первой фазе эякуляции благодаря общему укорочению семявыносящего протока, после чего он снова растягивается благодаря расслаблению продольных мышц до первоначальной длины и его просвет снова заполняется. В качестве резервуаров для накопления спермиев рассматривают хвост придатка и ампулярную часть семявыносящего протока [171, 535, 186, 472, 316, 420, 84].
Данные Лоде [435] и Экснера [356] об эякуляторной способности и наличии спермиев при повторных эякуляциях подвержены чрезвычайно широким видовым различиям. Если макака способна к четырем эякуляциям в сутки на протяжении 3—4 дней [324], а павиан — к трем эякуляциям в течение 20 мин [315], то золотистый хомячок в течение часа может произвести до 50 эякуляций [327]. Маккензи и Берлинер, собрав в течение суток 20 эякулятов барана, нашли, что 19-й эякулят содержал более миллиона спермиев при общем объеме 0,66 мл по сравнению с 3,5 миллионами спермиев при объеме 0,7 мл в первом эякуляте [451]. Количество извергаемого при одной садке эякулята варьирует от 0,1 —1,25 мл у козла до 150—500 мл у хряка [298].
Ван Дюзен, Слотер и Уайт [523], произведя в эксперименте на "собаках двустороннюю пресакральную симпатэктомию, продемонстрировали утрату эякуляции при сохранной эрекции. Симаяс и Лзпгуорси [49] на основании своих экспериментов на лотах пришли к выводу, что активная фаза эрекции представляет феномен чисто парасимпатический, в то время как эякуляция сочетает и симпатические, и парасимпатические реакции (рис. 34).
Изучение шерваторных (механизмов семявыносящего протока, семенных. пузырьков и простаты с акцентом на действие медиаторов проведено Шестрандом (Ш55). В частности, им осуществлено флюорометрическое исследование содержания катехоламинов в семявыносящих протоках и добавочных половых железах различных животных. Наряду с очень высоким содержанием норадреналина в исследованных анатомических образованиях автор обнаружил прямую корреляцию уровней норадреналина с количеством гладких мышц в этих органах: •органы с высоким содержанием железистой ткани выявляют меньшую концентрацию норадреналина, чем органы с выраженным мышечным слоем [505].
Психосенсорный эквивалент первой фазы эякуляции описан Мастерсом и Джонсон [449]: в отличие от острого переживания оргазма, синхронного с энергичными мышечными спазмами по
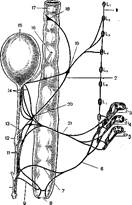
Рис. 34. Схема иннервации половых органов кота [496].
1 — симпатический ствол; ослабление эрекции; сокращение т. ге1гас1ог решз; сжатие сосудов; эякуляция; 2 — п. Ьуро-§азШсиз; ослабление эрекции; сокращение т. ге1гас1ог решз; и кровеносных сосудов; эякуляция; 3 — очень слабая эрекция; 4 — полная эрекция; эякуляция; мочеиспускание; 5— мочеиспускание; частичная эрекция; 6 — п. рийепйш 1п1.; прекращение эрекции; сокращение т. ге1гаЫх>г решз; эякуляция; 7 — п. йогзаНз ре-П18; 8— апиз; 9— полная эрекция; 10 — решз; 11 —нижняя, часть р!ехиз Ьуро§аз(;псиз; неполная эрекция; 12 — простата; 13 — средняя часть. р!ехиз пуроеавШсиз; эякуляция; прекращение эрекции; 14 — верхняя часть р!ехиз Ь>-роеазШсиз; сокращение мочевого пузыря; 15 — мочевой пузырь; 16 — иге1ег; 17 — гес-1ит; 18—2'1"Ь'1- соенасит; 1У— нижний брыжеечный узел;
перечно-полосатых мышц, извергающих эякулят и коррелирующих со второй фазой эякуляции, в первой фазе оргазма как такового еще нет, а возникает лишь ощущение его неотвратимости. Этот момент наступает за 2—4 с до выброса эякулят» Боре и Комарр [317], исследовав 529 больных с травматическими повреждениями спинного мозга и (или) конского хвоста,, констатировали сохранение эрекций при центральном параличе у 85—90% больных, а при периферическом — у 25% больных. Эякуляции сохранялись при центральном параличе у 8% больных и при периферическом — у 19% больных. При этом был отмечен парадоксальный факт: частота наступления беременности у жен больных с периферическим параличом превышала таковую при центральном параличе. Исследователи объяснили это прохождением церебральных импульсов через сохранившиеся пп. вр1апспп1с1, Манроу, Хорни и Полл [464] наблюдали 84 бывших военнослужащих с последствиями ранений позвоночника; некоторымТаблица 3
Частота нарушений эрекции и эякуляции при различных локализациях
симпатэктодгаи [532].
| Уровни | Общее число наблюдений | Нарушение эрекции | Стойкая утрата эякуляции |
| Грудной | |||
| Пояснично-грудной | |||
| Поясничный: | |||
| односторонние | |||
| двусторонние |
из них для снятия болезненной спастичности было произведено пересечение передних корешков. Сохранность основных сексуальных феноменов у этих больных оказалась близкой к данным Борса и Комарра: эрекции были_у 74% больных, а эякуляции— у 7—8%. Авторы приходят к выводу, что наличие или отсутствие расстройств потенции определяется не только разрушением спинного мозга как таковым, но и его локализацией и числом пораженных сегментов, так как, в частности, «ни в поясничном отделе спинного мозга, ни в каком-либо другом месте никакого „центра" эякуляции нет» [464]. Этот вывод Манроу, Хорни и Полла подтверждается данными Уайтло и Смисуика, изучавших влияние симпатэктомии на половые функции. Наряду с улучшением сексуальных проявлений, отмеченным 11 оперированными из 183 (7%), у 169 больных имели место жалобы па нарушение половых функций (табл. 3).
Объясняя наблюдавшиеся при этом нарушения •• эякуляции, авторы пишут: «Сам процесс выведения семенной жидкости при оперативных вмешательствах на симпатической нервной системе не страдает, но изгнание семени через член утрачивается. Парасимпатическая функция не изменяется. Количество сперматозоидов уменьшается, а оставшиеся не движутся из-за отсутствия мышечной активности. Вследствие нарушения сократительной способности гладких мышц семенных пузырьков в простатическую часть уретры между внутренним и наружным сфинктером поступает меньше семенной жидкости. Вследствие же расслабления внутреннего сфинктера и эта семенная жидкость, вместо того чтобы быть изгнанной наружу, в сторону члена, попадает в мочевой пузырь».
Основываясь на клинических наблюдениях, Уайтло и Смисуик предлагают следующие схемы инервационного обеспечения эрекции и эякуляции (пункты, противоречащие общепринятым взглядам и не подкрепленные собственной аргументацией, обозначены курсивом):



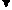

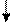








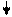
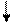














При всех недостатках и неточностях эти схемы все же дают наглядное представление о структурах, принимающих участие-в обеспечении эрекции и эякуляции, расположенных на различных уровнях, от ганглиозных клеток приорганных [456] тазовых сплетений до высших отделов головного мозга [193]. И хотя эякуляция непроизвольна, влияние на нее корковой регуляции прослеживается как при наступлении эякуляции под влиянием чисто психического возбуждения, без воздействия на эрогенную



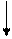

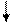







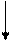




зону члена, так и при кортикально обусловленном функциональном асперматизме, когда коитус с определенной женщиной никогда не заканчивается эякуляцией, независимо от длительности фрикций. В клинико-физиологическом анализе механизмов кортикального обеспечения половых функций особого упоминания заслуживают парацентральные дольки, синдром поражения которых [53] свидетельствует о непосредственном их участии у человека в регулировке феномена эякуляции.
Оргазм
Оргазм (греч. оргазтоз, от огаб — разбухаю, пылаю страстью) — высшая степень сладострастного ощущения, возникающая в момент завершения полового акта (или заменяющих его <рорм половой активности).
Если нейрофизиологические механизмы эрекции и эякуляциш при неизбежных расхождениях между различными авторами по отдельным частным пунктам в общем вырисовываются в~ достаточно четкую и принимаемую большинством единую концепцию, то механизмы оргазма вое еще трактуются крайне противоречиво. Большое число гипотез, объясняющих природу оргазма, можно сгруппировать следующим образом:
I. Гипотезы периферического происхождения оргазма
1. Формирование оргазма связывается с сокращением семя-
извергающих мышц [350, 292].
2. Формирование оргазма связывается с активным преодолением семенной жидкостью узких устьев семявыносящих протоков, открывающихся в заднюю уретру на семенном бугорке. Сторонники этой гипотезы [480, 471] в свое время оперировали наиболее аргументированными доводами.. Например, Орловский пишет: «...нужно обратить внимание на то, что именно первая часть эякуляции вызывает оргазм, а не вторая. Прохождение в заднюю уретру через узкие выбрасывающие протоки содержимого семенных пузырьков, а также простатического сока вызывает чувство оргазма. Это может быть также ясно доказано впрыскиванием в заднюю уретру через гюйоновский наконечник 2% раствора кокаина перед половым актом... Произведенный затем обычным путем половой акт протекает без оргазма: прохождение через анестезированные отверстия не воспринимается организмом в виде особого ощущения. Это доказывается и кли